Résumé : L’utilisation de l’acupuncture de plus en plus fréquente dans l’assistance médicale à la procréation (AMP), surtout dans la fécondation in vitro (FIV) a engendré de nombreuses recherches pour tenter d’expliquer les mécanismes d’action dans l’infertilité féminine. La régulation de l’axe hypothalamo-hypophyso-ovarien et surrénalien, l’action sur le système nerveux sympathique et le flux sanguin ovarien sont quelques-uns des ces mécanismes physiopathologiques qui permettront de comprendre son action lors d’une dysfonction ovarienne. L’action sur le flux vasculaire utérin, le stress, les catécholamines, l’inhibition de la motilité utérine etc. sont d’autres mécanismes intervenant lors de la FIV avant et après transfert embryonnaire. Mots-clés : Acupuncture expérimentale – FIV – flux sanguin ovarien – leptine – VEGF – endothéline – NPY –connexine 43 – cytokines.
Summary: The use of acupuncture increasingly common in assisted reproductive technology (ART), especially in vitro fertilization (IVF) has created a lot of research to try to explain the mechanisms of action in female infertility. The regulation of the hypothalamic-pituitary-ovarian and adrenal action on the sympathetic nervous system and ovarian blood flow are some of the physiopathological mechanisms for understanding its action in an ovarian dysfunction. The action on the uterine vascular flow, stress, catecholamines, the inhibition of uterine motility etc. are other mechanisms involved in the before and after IVF embryo transfer. Keywords: Experimental Acupuncture – IVF – Ovarian blood flow – leptin – VEGF – endothelin – NPY-connexin 43 – cytokines.
De nombreuses théories visent à expliquer les mécanismes physiopathologiques de l’action de l’acupuncture dans l’infertilité. On peut les regrouper en quatre mécanismes principaux étroitement liés : la modulation des facteurs neuroendocriniens, l’augmentation du flux sanguin ovarien et utérin, la modulation du système immunitaire (en particulier les cytokines), la réduction du stress, de l’anxiété et de la dépression [1-4]. Ces mécanismes jouent un rôle potentiel aussi bien dans l’infertilité que dans l’explication des effets de l’acupuncture lors d’un protocole de FIV avant ou après transfert de l’embryon.

Mécanismes physiologiques dans l’infertilité en rapport avec une dysfonction ovarienne
Régulation de l’axe hypothalamo-hypophyso-ovarien et surrénalien
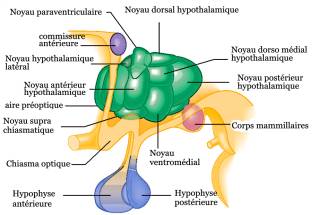
L’augmentation des bêta-endorphines engendrée par l’acupuncture ou l’électroacupuncture (EA) a un impact sur l’axe hypothalamo-hypophyso-ovarien mais aussi surrénalien en modulant la sécrétion des hormones GnRH et CRH (corticotropin-releasing hormone) chez la rate ovariectomisée ou présentant un syndrome d’ovaires polykystiques induits [5-9] ou chez la femme souffrant d’ovaires polykystiques [[10],[11]]. On observe ainsi que l’EA à basse fréquence entraîne une diminution de l’expression de la GnRH au niveau de l’aire médiale préoptique hypothalamique ainsi que de la LH dans l’hypophyse. Par ailleurs elle va aussi augmenter la synthèse de bêta-endorphines de nature hypothalamo-hypophysaire et plasmatique [6] et la libération de CRH dans ce même noyau paraventriculaire chez la rate ovariectomisée [9] et permettre la modulation du cortisol [[12]].
En outre de nombreuses investigations ont démontré que l’acupunture, la moxibustion ou l’EA à basse fréquence influencent les niveaux de FSH, LH, estradiol (E2) et progestérone [5,9,13-18]. Il s’avère que par exemple l’augmentation des œstrogènes plasmatiques par EA est liée à l’aromatisation extraglandulaire des androgènes [14], mais aussi par l’inhibition de la CRH d’origine ovarienne [2].
Action sur le flux sanguin ovarien : modulation par le système nerveux sympathique
L’acupuncture augmente le flux sanguin ovarien comme cela a été démontré par la stimulation EA à basse fréquence chez des rates avec ovaires polykystiques stéroïdes-induits. De nombreux travaux ont mis en évidence le rôle crucial du système nerveux sympathique dans le flux sanguin ovarien [2]. Après EA (2 Hz), l’activité du système sympathique décroit et le flux sanguin ovarien augmente, action engendrée via une réponse réflexe des nerfs sympathiques ovariens eux même régulés par les voies supraspinales [[19],[20]]. L’inhibition du système nerveux sympathique par EA (2Hz) sur des aiguilles placées au niveau de l’abdomen ou de la hanche de l’animal, correspond ainsi au territoire d’innervation de l’ovaire ou de l’utérus [[21]]. Chez la femme souffrant d’un syndrome des ovaires polykystiques (SOPK), l’EA à basse fréquence (2Hz) entraîne également une réduction de l’activité du système sympathique [[22]]. Par ailleurs, il a été démontré que l’EA (2Hz) module l’activité du système nerveux sympathique dans les ovaires en entraînant une décroissance de la concentration ovarienne de l’endothéline-1, de CRH et de NGF (nerve growth factor) [8,23-25], d’où régulation du flux sanguin ovarien par vasodilatation. Manni et coll. démontrent que l’EA à 2Hz entraîne une décroissance de l’activité sympathique en diminuant l’expression des récepteurs adrénergiques β2 aussi bien que celle des récepteurs neurotrophiques p75 (nerve growth factor receptor p75NTR ou NGFR qui permet le développement des neurones adrénergiques) [[26]].
Modulation du système endocrinien glucidique
L’hyperinsulinisme ou une résistance à l’insuline serait une des causes du SOPK caractérisé par l’association d’une spanioménorrhée ou d’une aménorrhée, d’un problème de stérilité, d’un hirsutisme, d’une obésité etc. et que 50% des patients présentant un SOPK le présenterait [[27],[28]]. L’EA (100Hz ou 2Hz) ou l’acupuncture réduit le poids corporel et la prise alimentaire, augmente la sensibilité à l’insuline, réduit la glycémie et les niveaux de lipides, stimule aussi le transport du glucose dans le muscle squelettique aussi bien chez les rats [29-32] que l’être humain [33-35], le tout en rapport avec une modulation de la concentration plasmatique en leptine ou adiponectine [29,30,[36]]. L’acupuncture pourrait conduire à réduire le poids dans le SOPK et améliorer le cycle menstruel via la régulation de l’activité de la leptine et de l’axe hypothalamo-hypophyso-ovarien [[37]].
Mécanismes physiologiques impliqués lors de la FIV avant transfert embryonnaire
Action sur le VEGF, le flux vasculaire utérin, le flux sanguin endométrial
En cas d’hypoxie dans le fluide folliculaire, le vascular endothelial growth factor (VEGF) sera produit, facteur de l’angiogenèse. Il est ainsi connu que des concentrations élevées de VEGF dans ce liquide peuvent être une des causes d’échec de la FIV [[38]]. Or, l’acupuncture module la production du vascular endothelial growth factor (VEGF) [17,39-41] (figure 1).
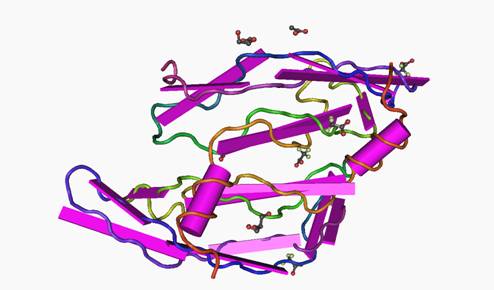
Figure 1. Structure cristalline de la VEGF-A, impliquée dans l’angiogenèse.
Avant le transfert de l’embryon, la pression artérielle et l’activité vasoconstrictive du système sympathique seront réduites par l’intermédiaire de l’action de l’acupuncture sur les bêta-endorphines et sur la régulation de l’axe hypothalamo-hypophyso-ovarien [2]. Ainsi après huit séances d’EA (à 2 et 100 Hz) réparties sur quatre semaines, l’impédance (résistance) vasculaire dans les artères utérines chez les femmes stériles traitées par analogue de GnRH fut réduite de manière statistiquement significative et maintenue quinze jours après la fin du traitement EA [[42]]. L’inhibition du système nerveux sympathique semble être en corrélation avec cette diminution de la résistance vasculaire utérine [42,[43]], elle même en rapport avec l’augmentation de la concentration en bêta-endorphines [44,45].
Sze So et coll. ont objectivé par contre que l’acupuncture réduisait le flux sanguin de l’endomètre mesuré par doppler entraînant une hypoxie endométriale qui améliore, elle, l’implantation de l’embryon [[46]].
Action sur le stress, l’anxiété et la dépression
L’association entre prévalence d’un syndrome dépressif et échec de FIV a été objectivée chez des femmes australiennes en traitement de stérilité [[47]] de même qu’avec l’anxiété et le stress [[48]]. Le tout suggère que le stress, l’anxiété et la dépression sont des composantes à traiter chez les femmes réalisant une FIV.
Ainsi les femmes ayant un échec de leur FIV ont une concentration en catécholamines (adrénaline et noradrénaline) supérieure à celles dont la FIV est une réussite [[49]]. On retrouve aussi une augmentation du cortisol et de la prolactine chez les femmes réalisant une FIV, non retrouvée chez celles qui bénéficient d’une laparoscopie sans relation avec une infertilité [[50]]. L’acupuncture montre que son efficacité à réduire le stress et l’anxiété à la fois avant et après le transfert d’embryon peut améliorer les taux de grossesse [[51]].
Le jour du transfert embryonnaire, l’acupuncture réduit aussi la concentration en cortisol et diminue l’anxiété [46]. Une autre étude observe que l’électroacupuncture augmente les taux de cortisol et de prolactine de manière significative, respectivement du 7ème au 13ème jour et du 5ème au 8ème jour après stimulation par agoniste de GnRH par rapport au groupe contrôle puis retourne à l’état basal physiologique au moment du transfert. Les auteurs concluent que l’acupuncture module les taux de prolactine et de cortisol de façon à ce que le corps retrouve son état homéostasique [12].
C’est en modulant les niveaux de neuropeptide Y (NPY) que l’acupuncture réduit la dépression, l’anxiété et le stress. Par exemple, le traitement acupunctural diminue l’anxiété comportementale chez les rats en augmentant la concentration du NPY dans l’amygdale [[52]]. De même, dans une groupe de femmes bénéficiant d’EA à 2Hz sur 5TR et 4GI, d’EA à 80Hz sur le 29E et d’acupuncture manuelle sur le 20VG et le 36E dans le cadre d’une analgésie pour aspiration ovocytaire lors d’une FIV, on observe une augmentation statistiquement significative (p<0,001) de la concentration du NPY dans le liquide folliculaire versus le groupe alfentanyl avec diminution du stress et de la prise d’antalgiques [[53]].
De nombreux mécanismes potentiels des effets de l’acupuncture ont été évoqués dans les troubles de l’humeur, l’anxiété et le stress [4,[54]]. Outre la modulation de l’augmentation du NPY, on retrouve aussi un accroissement de la concentration des peptides opioïdes [[55]], la restauration des taux de brain-derived neurotrophic factor (BDNF) au niveau de l’hippocampe [[56]], l’atténuation du système nerveux sympathique [[57]] et son corollaire, l’augmentation de l’activité du système vagal [[58]], la modulation de la prolactine [12] et bien sûr l’influence sur l’axe hypothalamo-hypophyso-surrénalien [[59]].
Action sur le système immunitaire
Il semble que les échecs répétés des FIV soient en rapport avec une modulation locale et systémique inappropriée de la réponse des lymphocytes T CD4+ helper à réponse Th2 [[60]]. La libération de cytokines tels que les interleukine-4 (IL-4), IL-6, IL10 et IL-13 est associée à la réponse des lymphocytes Th2 qui favorisent les réactions humorales de type allergique avec activation des cellules éosinophiles et des plasmocytes. La grossesse réussie a été décrite comme un « phénomène Th-2 ». En effet, les taux sériques significativement élevés de cytokines à réponse Th2 (IL-6 et IL-10) sont détectés dans la grossesse normale, alors que chez les femmes avec avortements récurrents, les taux sériques des cytokines à réponse Th1 et l’IFN-γ sont plus élevés [[61]]. L’acupuncture pourrait améliorer la FIV en agissant sur ces cytokines [[62]]. L’acupuncture, l’EA ou la moxibustion moduleraient les concentrations de cytokines issues des réponses Th1 à médiation cellulaire ou Th2 à médiation humorale [63-65]. Ce qui semble confirmé par deux études parues en janvier 2012 qui montrent que sur un modèle de rates en échec d’implantation du blastocyste (par ingestion de mifépristone), l’acupuncture va améliorer l’état déficient de la muqueuse utérine en promouvant la sécrétion des cytokines à réponse Th2 (IL-4, IL-10) et en inhibant les cytokines à réponse Th1 (IL-1β, IL-2) [[66]], mais aussi en améliorant la sécrétion d’IL-12 et de LIF (leukemia inhibitory factor) [[67]].
Mécanismes physiologiques impliqués lors de la FIV après transfert embryonnaire
Connexine 43
L’implantation du blastomère serait améliorée après puncture des points 3F, 36E et 6Rte dès le premier jour de grossesse chez des rates gravides. Une relation avec la connexine 43 a été mise en évidence [[68]]. Les connexines sont des protéines transmembranaires qui s’assemblent en complexes de six unités, le connexon qui va ménager ainsi une jonction percée par un pore reliant les cytoplasmes des deux cellules contiguës entre les membranes de deux cellules voisines (figure 2). D’où l’importance de ces structures qui permettent le passage des éléments nécessaires tout particulièrement chez l’embryon car assurant ainsi la circulation des nutriments en attendant la formation du système sanguin.
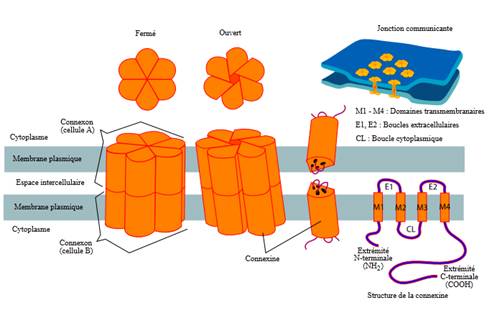
Figure 2. Jonction gap avec son principal élément, le connexon et la la connexine, protéine formant les connexons (Mariana Ruiz LadyofHats [Public domain], via Wikimedia Commons).
Motilité utérine
L’observation des contractions utérines de haute fréquence (> 5,0 contractions/min) au moment du transfert embryonnaire a été associée à une implantation et un taux de grossesse nettement plus faible par transfert d’embryon par comparaison avec les femmes ayant une fréquence de contractions plus faibles (≤ 3,0 contractions/min). Cela peut entraîner l’expulsion mécanique d’embryons à partir de la cavité utérine [[69]]. D’où l’intérêt de les réduire [[70]].
Ainsi la puncture de 4GI entraîne une réduction statistiquement significative de la motilité utérine chez les rates gravides. Ce serait en rapport avec le rôle inhibiteur de l’expression de l’enzyme COX-2 par inactivation des prostaglandines [[71]]. De même, le traitement acupunctural de 6Rte contrôle aussi la motilité utérine pendant la grossesse [[72]].
Action sur les facteurs de réceptivité de l’endomètre lors de l’implantation
La stimulation des points 6Rte, 36E, 3F, 4VC, 3VC entrainent une amélioration de l’expression de la protéine du facteur d’inhibition de l’endomètre (LIF) et de l’ostéopontine (OPN) au niveau de l’utérus gravide d’un modèle de rate induite par citrate de clomifène (correspond à un syndrome des ovaires polykystiques) pendant la période d’implantation du foetus. On sait que ces deux facteurs cellulaires sont considérés comme les biomarqueurs les plus prometteurs de la réceptivité de l’endomètre lors de l’implantation du blastocyste et durant la grossesse. Par ailleurs, les taux sériques d’estradiol sont diminués de manière significative [[73]].
Références
[1]. Stéphan JM. Acupuncture expérimentale dans l’insuffisance ovarienne. Acupuncture & Moxibustion. 2005;4(1):68-72.
[2]. Stéphan JM. Acupuncture expérimentale et syndrome des ovaires polykystiques. Acupuncture & Moxibustion. 2005;4(2):153-159.
[3]. Stéphan JM. Acupuncture expérimentale et grossesse. Acupuncture & Moxibustion. 2005;4(3):236-239.
[4]. Stéphan JM. Acupuncture expérimentale, stress, axe neuroendocrinien et système limbique. Acupuncture & Moxibustion. 2005;4(4):340-349.
[5]. Chen BY, Yu J. Relationship between blood radioimmunoreactive beta-endorphin and hand skin temperature during the electro-acupuncture induction of ovulation. Acupunct Electrother Res 1991;16(1-2):1-5.
[6]. Chen BY. Acupuncture normalizes dysfunction of hypothalamic-pituitary-ovarian axis. Acupunct Electrother Res 1997;22(2):97-108.
[7]. Stener-Victorin E, Lindholm C. Immunity and beta-endorphin concentrations in hypothalamus and plasma in rats with steroid-induced polycystic ovaries: effect of low-frequency electroacupuncture. Biol Reprod. 2004 Feb;70(2):329-33.
[8]. Stener-Victorin E, Lundeberg T, Waldenström U, Bileviciute-Ljungar I, Janson PO. Effects of electro-acupuncture on corticotropin-releasing factor in rats with experimentally-induced polycystic ovaries. Neuropeptides. 2001 Oct-Dec;35(5-6):227-31.
[9]. Zhao H, Tian ZZ, Chen BY. Increased corticortropin-releasing hormone release in ovariectomized rats’paraventricular nucleus: effects of electroacupuncture. Neurosci Lett 2003;353(1):37-40.
[10]. Stener-Victorin E, Waldenstrom U, Tagnfors U, Lundeberg T, Lindstedt G, Janson PO. Effects of electro-acupuncture on anovulation in women with polycystic ovary syndrome. Acta Obstet Gynecol Scand 2000 79:180-188.
[11]. Jedel E, Labrie F, Odén A, Holm G, Nilsson L, Janson PO, Lind AK, Ohlsson C,Stener-Victorin E. Impact of electro-acupuncture and physical exercise on hyperandrogenism and oligo/amenorrhea in women with polycystic ovary syndrome: a randomized controlled trial. Am J Physiol Endocrinol Metab. 2011 Jan;300(1):E37-45.
[12]. Magarelli PC, Cridennda DK, Cohen M. Changes in serum cortisol and prolactin associated with acupuncture during controlled ovarian hyperstimulation in women undergoing in vitro fertilization-embryo transfer treatment. Fertil Steril. 2009 Dec;92(6):1870-9.
[13]. Chen BY, Cheng LH, Gao H, Ji SZ. [Effects of electroacupuncture on the expression of estrogen receptor protein and mRNA in rat brain] Sheng Li Xue Bao 1998;50(5):495-500.
[14]. Zhao H, Tian Z, Cheng L, Chen B. Electroacupuncture enhances extragonadal aromatization in ovariectomized rats. Reprod Biol Endocrinol. 2004 Apr 27;2:18.
[15]. Liu F, Xiong J, Huang GY, Wang W. [Study on the underlying mechanism of acupuncture in regulating neuroendocrine activity in dysmenorrhea rats]. Zhen Ci Yan Jiu. 2009 Feb;34(1):3-8.
[16]. Yang MC, Han LJ, Li J, Fang G, Xiao XQ. [Effects of medicated thread moxibustion of Zhuang medicine on sex hormone in ovariectomized rabbits]. Zhongguo Zhen Jiu. 2011 Feb;31(2):145-8.
[17]. He DJ, Huang GY, Zhang MM. [Effects of acupuncture on the luteal function of rats with dysfunctional embryo implantation]. Zhongguo Zhen Jiu. 2009 Nov;29(11):910-3.
[18]. Chen J, Liu LL, Cui W, Sun W. [Effects of electroacupuncture on in vitro fertilization-embryo transfer (IVF-ET) of patients with poor ovarian response]. Zhongguo Zhen Jiu. 2009 Oct;29(10):775-9.
[19]. Stener-Victorin E, Kobayashi R, Kurosawa M. Ovarian blood flow responses to electro-acupuncture stimulation at different frequencies and intensities in anaesthetized rats. Auton Neurosci 2003;108(1-2):50-6.
[20]. Stener-Victorin E, Fujisawa S, Kurosawa M. Ovarian blood flow responses to electroacupuncture stimulation depend on estrous cycle and on site and frequency of stimulation in anesthetized rats. J Appl Physiol. 2006 Jul;101(1):84-91.
[21]. Stener-Victorin E, Kobayashi R, Watanabe O, Lundeberg T, and Kurosawa M. Effect of electro-acupuncture stimulation of different frequencies and intensities on ovarian blood flow in anaesthetised rats with steroid-induced polycystic ovaries. Reprod Biol Endocrinol. 2004;2:16.
[22]. Stener-Victorin E, Jedel E, Janson PO, Sverrisdottir YB. Low-frequency electroacupuncture and physical exercise decrease high muscle sympathetic nerve activity in polycystic ovary syndrome. Am J Physiol Regul Integr Comp Physiol. 2009 Aug;297(2):R387-95.
[23]. Stener-Victorin E, Lundeberg T, Cajander S, Aloe L, Manni L, Waldenstrom U, Janson PO. Steroid-induced polycystic ovaries in rats: effect of electro-acupuncture on concentrations of endothelin-1 and nerve growth factor (NGF), and expression of NGF mRNA in the ovaries, the adrenal glands, and the central nervous system.Reprod Biol Endocrinol 2003;1(1):33.
[24]. Stener-Victorin E, Lundeberg T, Waldenstrom U, Manni L, Aloe L, Gunnarsson S, Janson PO. Effects of electro-acupuncture on nerve growth factor and ovarian morphology in rats with experimentally induced polycystic ovaries. Biol Reprod 2000;63(5):1497-503.
[25]. Bai YH, Lim SC, Song CH, Bae CS, Jin CS, Choi BC, Jang CH, Lee SH, Pak SC. Electro-acupuncture reverses nerve growth factor abundance in experimental polycystic ovaries in the rat. Gynecol Obstet Invest 2004;57(2):80-5.
[26]. Manni L, Lundeberg T, Holmäng A, Aloe L, Stener-Victorin E. Effect of electro-acupuncture on ovarian expression of alpha (1)- and beta (2)-adrenoceptors, and p75 neurotrophin receptors in rats with steroid-induced polycystic ovaries. Reprod Biol Endocrinol. 2005 Jun 7;3:21.
[27]. Hopkinson ZE, Sattar N, Fleming R, Greer IA. Polycystic ovary syndrome: the metabolic syndrome comes to gynaecology. Br Med 1998;J 317:329332 .
[28]. Legro RS, Kunselman AR, Dodson WC, Dunaif A. Prevalence and predictors of risk for type 2 diabetes mellitus and impaired glucose tolerance in polycystic ovary syndrome: a prospective, controlled study in 254 affected women. J Clin Endocrinol Metab. 1999 Jan;84(1):165-9.
[29]. Kim SK, Lee G, Shin M, Han JB, Moon HJ, Park JH, Kim KJ, Ha J, Park DS, Min BI. The association of serum leptin with the reduction of food intake and body weight during electroacupuncture in rats. Pharmacol Biochem Behav. 2006 Jan;83(1):145-9.
[30]. Mannerås L, Jonsdottir IH, Holmäng A, Lönn M, Stener-Victorin E. Low-frequency electro-acupuncture and physical exercise improve metabolic disturbances and modulate gene expression in adipose tissue in rats with dihydrotestosterone-induced polycystic ovary syndrome. Endocrinology. 2008 Jul;149(7):3559-68.
[31]. Johansson J, Feng Y, Shao R, Lönn M, Billig H, Stener-Victorin E. Intense electroacupuncture normalizes insulin sensitivity, increases muscle GLUT4 content, and improves lipid profile in a rat model of polycystic ovary syndrome. Am J Physiol Endocrinol Metab. 2010 Oct;299(4)
[32] . Wang SJ, Xu HZ, Xiao HL. [Effect of high-frequency electroacupuncture on lipid metabolism in obesity rats]. Zhen Ci Yan Jiu. 2008 Jun;33(3):154-8.
[33]. Cabioglu MT, Gündogan N, Ergene N. The efficacy of electroacupuncture therapy for weight loss changes plasma lipoprotein A, apolipoprotein A and apolipoprotein B levels in obese women. Am J Chin Med. 2008;36(6):1029-39.
[34] . Cabioğlu MT, Ergene N. Changes in levels of serum insulin, C-Peptide and glucose after electroacupuncture and diet therapy in obese women. Am J Chin Med. 2006;34(3):367-76.
[35]. Cabioğlu MT, Ergene N. Changes in serum leptin and beta endorphin levels with weight loss by electroacupuncture and diet restriction in obesity treatment. Am J Chin Med. 2006;34(1):1-11.
[36]. Luo HL, Li RH. [Effect of electroacupuncture on leptin and adiponectin in simple obesity patients]. Zhen Ci Yan Jiu. 2007 Aug;32(4):264-7.
[37]. Li MY, Wei DY, Wu JM. [Analysis on the interrelation between acupuncture-induced weight reduction and menstruation]. Zhen Ci Yan Jiu. 2007 Apr;32(2):142-4.
[38]. Barroso G, Barrionuevo M, Rao P, Graham L, Danforth D, Huey S, Abuhamad A, Oehninger S. Vascular endothelial growth factor, nitric oxide, and leptin follicular fluid levels correlate negatively with embryo quality in IVF patients. Fertil Steril. 1999 Dec;72(6):1024-6.
[39]. Zhu CJ, Dong JX, Zhang MJ, Lu GL, Li J. [Effect of acupoint injection with bone marrow mesenchymal stem cells on the blood flow in rats with hind limb ischemia]. Zhongguo Zhen Jiu. 2009 Dec;29(12):987-92.
[40]. Zhang DS, Lu YK, Su ZR, Cai DK. [Effect of acupoint sticking of « Hua yutie » on VEGF expression in rats of focal cerebral ischemia]. Zhongguo Zhen Jiu. 2009 Mar;29(3):217-21.
[41]. Ma J, Luo Y. Effects of electroacupuncture on expressions of angiogenesis factors and anti-angiogenesis factors in brain of experimental cerebral ischemic rats after reperfusion. J Tradit Chin Med. 2008 Sep;28(3):217-22.
[42]. Stener-Victorin E, Waldenstrom U, Andersson SA, Wikland M. Reduction of blood flow impedance in the uterine arteries of infertile women with electro-acupuncture. Hum Reprod 1996;11(6):1314-7.
[43]. Ho M, Huang LC, Chang YY, Chen HY, Chang WC, Yang TC, Tsai HD. Electroacupuncture reduces uterine artery blood flow impedance in infertile women. Taiwan J Obstet Gynecol. 2009 Jun;48(2):148-51.
[44]. Ku YH, Chang YZ. Beta-endorphin- and GABA-mediated depressor effect of specific electroacupuncture surpasses pressor response of emotional circuit. Peptides. 2001 Sep;22(9):1465-70.
[45]. Han JS. Acupuncture and endorphins. Neurosci Lett. 2004 May 6;361(1-3):258-61.
[46]. So EW, Ng EH, Wong YY, Lau EY, Yeung WS, Ho PC. A randomized double blind comparison of real and placebo acupuncture in IVF treatment. Hum Reprod. 2009 Feb;24(2):341-8.
[47]. Thiering P, Beaurepaire J, Jones M, Saunders D, Tennant C. Mood state as a predictor of treatment outcome after in vitro fertilization/embryo transfer technology (IVF/ET). J Psychosom Res. 1993 Jul;37(5):481-91.
[48]. Klonoff-Cohen H, Natarajan L, Klonoff E. Validation of a new scale for measuring Concerns of Women Undergoing Assisted Reproductive Technologies (CART). J Health Psychol. 2007 Mar;12(2):352-6.
[49]. Smeenk JM, Verhaak CM, Vingerhoets AJ, Sweep CG, Merkus JM, Willemsen SJ, van Minnen A, Straatman H, Braat DD. Stress and outcome success in IVF: the role of self-reports and endocrine variables. Hum Reprod. 2005 Apr;20(4):991-6.
[50]. Harlow CR, Fahy UM, Talbot WM, Wardle PG, Hull MG. Stress and stress-related hormones during in-vitro fertilization treatment. Hum Reprod. 1996 Feb;11(2):274-9.
[51]. Balk J, Catov J, Horn B, Gecsi K, Wakim A. The relationship between perceived stress, acupuncture, and pregnancy rates among IVF patients: a pilot study. Complement Ther Clin Pract. 2010 Aug;16(3):154-7.
[52] . Park HJ, Chae Y, Jang J, Shim I, Lee H, Lim S. The effect of acupuncture on anxiety and neuropeptide Y expression in the basolateral amygdala of maternally separated rats. Neurosci Lett 2005;377(3):179-84.
[53]. Stener-Victorin E, Waldenström U, Wikland M, Nilsson L, Hägglund L, Lundeberg T. Electro-acupuncture as a peroperative analgesic method and its effects on implantation rate and neuropeptide Y concentrations in follicular fluid. Hum Reprod. 2003 Jul;18(7):1454-60.
[54]. Stéphan JM. Acupuncture expérimentale, stress et molécules informationnelles. Acupuncture & Moxibustion. 2006;5(2):162-170.
[55]. Han SH, Yoon SH, Cho YW, Kim CJ, Min BI. Inhibitory effects of electroacupuncture on stress responses evoked by tooth-pulp stimulation in rats. Physiol Behav. 1999 Apr;66(2):217-22.
[56]. Yun SJ, Park HJ, Yeom MJ, Hahm DH, Lee HJ, Lee EH. Effect of electroacupuncture on the stress-induced changes in brain-derived neurotrophic factor expression in rat hippocampus. Neurosci Lett 2002;318(2):85-8.
[57]. Middlekauff HR, Hui K, Yu JL, Hamilton MA, Fonarow GC, Moriguchi J, et al. Acupuncture inhibits sympathetic activation during mental stress in advanced heart failure patients. J Card Fail 2002;8(6):399-406.
[58]. Wang JD, Kuo TB, Yang CC. An alternative method to enhance vagal activities and suppress sympathetic activities in humans. Auton Neurosci. 2002 Sep 30;100(1-2):90-5.
[59]. Cui H, Xiahong L, Xuewu L et Al. [Effects of puncturing the acupoints of baihui and sanyinjiao by electric needles on the HPA axis in the rat chronic stress-induced depression model]. Journal of Beijing University of traditional chinese medecine 2001;24(3):74.
[60]. Bermas BL, Hill JA. Proliferative responses to recall antigens are associated with pregnancy outcome in women with a history of recurrent spontaneous abortion. J Clin Invest. 1997 Sep 15;100(6):1330-4. Erratum in: J Clin Invest 1998 Jan 15;101(2):513.
[61]. Raghupathy R, Makhseed M, Azizieh F, Hassan N, Al-Azemi M, Al-Shamali E. Maternal Th1- and Th2-type reactivity to placental antigens in normal human pregnancy and unexplained recurrent spontaneous abortions. Cell Immunol. 1999 Sep 15;196(2):122-30.
[62]. Anderson BJ, Haimovici F, Ginsburg ES, Schust DJ, Wayne PM. In vitro fertilization and acupuncture: clinical efficacy and mechanistic basis. Altern Ther Health Med. 2007 May-Jun;13(3):38-48.
[63]. Gao M, Wu Y, Li GQ. [Regulating effect of moxibustion pretreatment on Th1/Th2 imbalance of athletes during course of heavy load training]. Zhongguo Zhen Jiu. 2011 Mar;31(3):247-51.
[64]. Liu XY, Zhou HF, Pan YL, Liang XB, Niu DB, Xue B, Li FQ, He QH, Wang XH, Wang XM. Electro-acupuncture stimulation protects dopaminergic neurons from inflammation-mediated damage in medial forebrain bundle-transected rats. Exp Neurol. 2004 Sep;189(1):189-96.
[65]. Ouyang BS, Che JL, Gao J, Zhang Y, Li J, Yang HZ, Hu TY, Wu YJ, Yang M. [Effects of electroacupuncture and simple acupuncture on changes of IL-1, IL-4, IL-6 and IL-10 in peripheral blood and joint fluid in patients with rheumatoid arthritis]. Zhongguo Zhen Jiu. 2010 Oct;30(10):840-4.
[66]. Gui J, Xiong F, Li J, Huang G. Effects of acupuncture on Th1, th2 cytokines in rats of implantation failure. Evid Based Complement Alternat Med. 2012;2012:893023. Epub 2012 Jan 23.
[67]. Gui J, Xiong F, Li J, Huang G. Effects of Acupuncture on LIF and IL-12 in Rats of Implantation Failure. Am J Reprod Immunol. 2012 Jan 9.
[68]. Huang GY, Zheng CH, Wu YX, Wang W. Involvement of connexin 43 in the acupuncture effect of improving rat blastocyst implantation. Fertil Steril. 2010 Mar 15;93(5):1715-7.
[69]. Fanchin R, Righini C, Olivennes F, Taylor S, de Ziegler D, Frydman R. Uterine contractions at the time of embryo transfer alter pregnancy rates after in-vitro fertilization. Hum Reprod. 1998 Jul;13(7):1968-74.
[70]. Fanchin R, Ayoubi JM, Righini C, Olivennes F, Schönauer LM, Frydman R. Uterine contractility decreases at the time of blastocyst transfers. Hum Reprod. 2001 Jun;16(6):1115-9.
[71]. Kim J, Shin KH, Na CS. Effect of acupuncture treatment on uterine motility and cyclooxygenase-2 expression in pregnant rats. Gynecol Obstet Invest 2000;50(4):225-30.
[72]. Kim JS, Na CS, Hwang WJ, Lee BC, Shin KH, Pak SC. Immunohistochemical localization of cyclooxygenase-2 in pregnant rat uterus by Sp-6 acupuncture. Am J Chin Med 2003;31(3):481-8.
[73]. Fu H, He Y, Gao Y, Man Y, Liu W, Hao H. Acupuncture on the Endometrial Morphology, the Serum Estradiol and Progesterone Levels, and the Expression of Endometrial Leukaemia-inhibitor Factor and Osteopontin in Rats. Evid Based Complement Alternat Med. 2011;2011:606514.

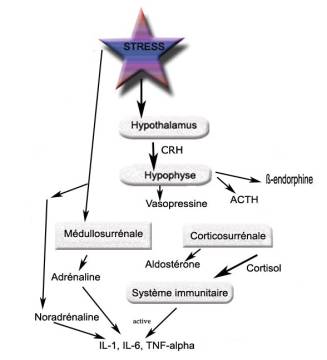
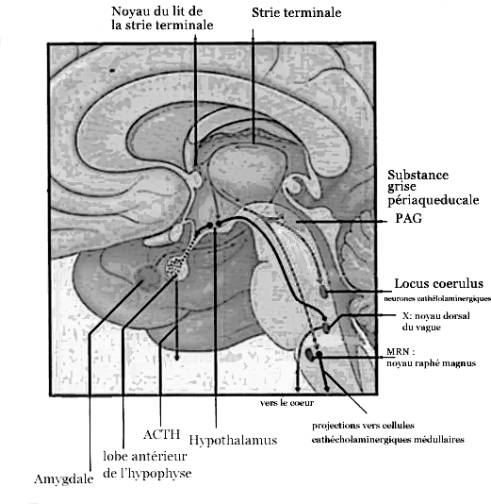
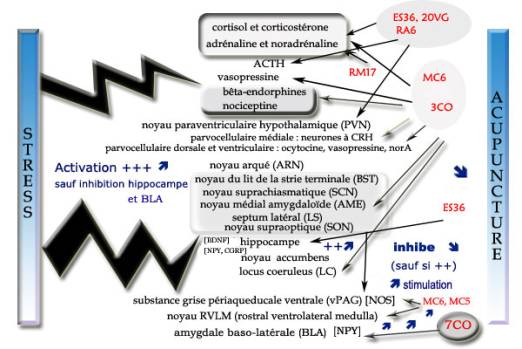
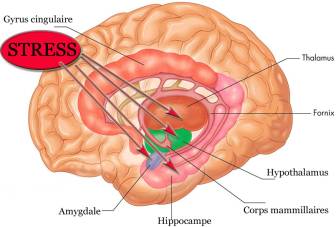 Figure 2. Principales structures intervenant dans la réponse au stress.
Figure 2. Principales structures intervenant dans la réponse au stress.