Résumé : La douleur neuropathique résulte d’un mécanisme de sensibilisation périphérique et centrale, modulée par les contrôles inhibiteurs spinaux et supraspinaux. Comme le montrent les nombreuses études acupuncturales expérimentales sur modèle animal de neuropathie, le contrôle de la douleur peut s’effectuer par modulation de différentes molécules et récepteurs comme le GABA, le glutamate et ses différents récepteurs (AMPA, NMDA…), le système sérotoninergique (5HT), le système opioïde, la noradrénaline (NE), les récepteurs cholinergiques etc. On note que sur les douleurs neuropathiques, l’électroacupuncture (EA) à basse fréquence (2Hz) a une action analgésique bien supérieure à celle à haute fréquence (100 Hz).
Summary: Neuropathic pain results from a mechanism of peripheral and central sensitization, modulated by spinal and supraspinal controls inhibitors. As shown in many acupuncture experimental studies of animal model of neuropathy, pain control may be effected by modulation of different molecules and receptors such as GABA, glutamate and its different receptors (AMPA, NMDA …), the serotonin (5HT) system, the opioid system, norepinephrine (NE), cholinergic receptors etc. Note that on neuropathic pain, electroacupuncture (EA) at low frequency (2 Hz) has analgesic effect much higher than high-frequency (100 Hz).
La douleur neuropathique résulte d’une sensibilisation périphérique, centrale et d’une plasticité synaptique au niveau du système nerveux central (SNC) impliquant de nombreuses molécules que l’acupuncture module au niveau du neurone, voire localement au niveau du tissu cutané [1]. Cette algie, qu’elle soit neuropathique ou même nociceptive, suscite nécessairement la mise en jeu de contrôles inhibiteurs exercés par des structures aussi bien spinales que supraspinales.
Les axones des neurones nociceptifs de la corne postérieure constituent ainsi les faisceaux médullaires ascendants des voies extra-lemniscales. Ceux-ci projettent leur information à différents niveaux supraspinaux : thalamus (noyau ventral postéro-latéral VPL), substance grise périaqueducale, noyau cunéiforme, hypothalamus, système limbique avec le complexe amygdalien, puis projection vers les aires somesthésiques S1 et S2 du cortex pariétal. Certains neurones du thalamus médian projettent aussi leurs axones vers les aires corticales frontales, le cortex insulaire et le cortex cingulaire antérieur, impliqués dans les réactions émotionnelles plus élaborées à la douleur [2].
La douleur neuropathique, quant à elle entraîne des changements d’activité des réseaux thalamo-corticaux, qui développent des processus autonomes qui entretiennent la douleur. La représentation corticale des territoires corporels change après une lésion nerveuse. Ces changements pourraient être corrélés à la survenue des douleurs de type neuropathique.
Le cortex cingulaire antérieur (CCA) est impliqué ainsi de façon majeure dans cette perception. Les neurones du CCA sont des neurones inhibiteurs produisant du GABA et/ou neuropeptides recevant souvent une innervation glutamatergique. Le glutamate est le principal transmetteur excitateur rapide dans le cortex cingulaire antérieur (CCA). Différents types de récepteurs du glutamate (AMPA, KA, NMDA) ainsi que des récepteurs métabotropiques liés à une protéine G (mGluR1 et mGluR5) y sont distribués et engagés dans la réponse excitatrice à l’origine de l’algie neuropathique. Par ailleurs, on notera aussi l’implication du cortex insulaire, structure aussi du système limbique [3,4].

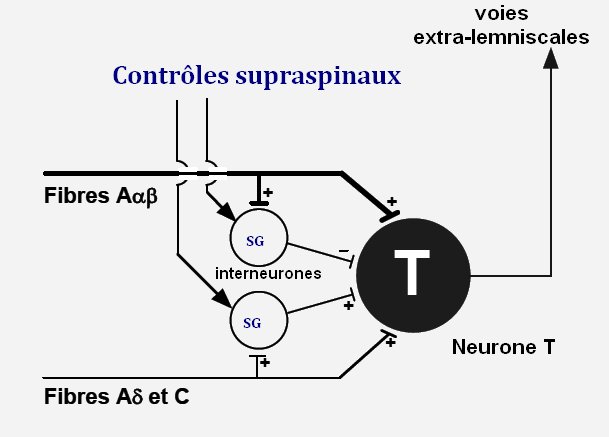
Figure 1. Contrôle segmentaire du « Gate control » (théorie du portillon de Melzack & Wall).
Les mécanismes de contrôle de la douleur : supraspinaux, structures cérébrales et cortex cingulaire antérieur
De manière schématique, on distingue quatre catégories de systèmes de contrôle.
Les contrôles segmentaires spinaux
La mise en jeu du contrôle segmentaire a été modélisée par Melzack et Wall [4] dans leur « théorie du portillon » (gate control theory) [5,6]. Ils ont mis en évidence l’importance du rôle de la corne postérieure de la moelle épinière dans la modulation de la transmission des messages nociceptifs. Ce modèle repose sur l’équilibre d’une balance entre deux types d’activités exercées sur les neurones nociceptifs non spécifiques médullaires. Les fibres nociceptives de petit diamètre Aδ et C d’origine segmentaire périphérique sont activatrices alors que les autres fibres de gros diamètre non nociceptives Aα, β de même origine périphérique sont inhibitrices. L’activation des fibres de gros diamètre de la sensibilité tactile légère Aα, β inhibe ainsi les réponses nociceptives par des mécanismes d’inhibition exercés par l’intermédiaire de l’activation de deux familles d’interneurones segmentaires localisés dans la couche II (substance gélatineuse). L’une est inhibitrice des neurones à convergence à l’origine des faisceaux ascendants des voies extra-lemniscales (neurone T « trigger cells ») activés par les fibres Aα, β et l’autre activatrice des neurones T activés par les fibres Aδ et C. Ces deux familles sont sous le contrôle de systèmes descendants d’origine supraspinale par l’intermédiaire d’acides aminés inhibiteurs comme la glycine ou l’acide gamma aminobutyrique (GABA). L’activation des fibres de la sensibilité tactile légère Aα, β, en augmentant l’activité des interneurones inhibiteurs, fermerait le portillon et bloquerait la transmission de l’information nociceptive vers les structures supraspinales (figure 1).
Les contrôles inhibiteurs descendants supraspinaux
Les contrôles inhibiteurs descendants issus du tronc cérébral s’exercent principalement à partir de deux structures à l’origine de voies descendantes dans la moelle épinière, l’une mésencéphalique avec la substance grise périaqueducale (SGPA), et l’autre bulbaire avec la région bulbaire ventromédiane (RBVM) associant le noyau du raphé magnus (NRM) et les noyaux paragigantocellulaire et gigantocellulaire. La stimulation électrique de ces structures entraîne des effets analgésiques résultant de la mise en jeu de voies descendantes sérotoninergiques exerçant un contrôle inhibiteur sur les neurones nociceptifs non spécifiques médullaires par libération de sérotonine et d’opioïdes endogènes dans les couches superficielles de la corne postérieure [7]. Il existe aussi une boucle de rétroaction négative spinobulbospinale mise en jeu par les axones des neurones sérotoninergiques qui se projettent directement dans la corne postérieure de la moelle. Par ailleurs, on a également la mise en jeu de systèmes inhibiteurs descendants noradrénergiques issus du locus coeruleus et du locus subcoeruleus en rapport avec des récepteurs α2-noradrénergiques activés physiologiquement par la noradrénaline (figure 2) [8,2 ].
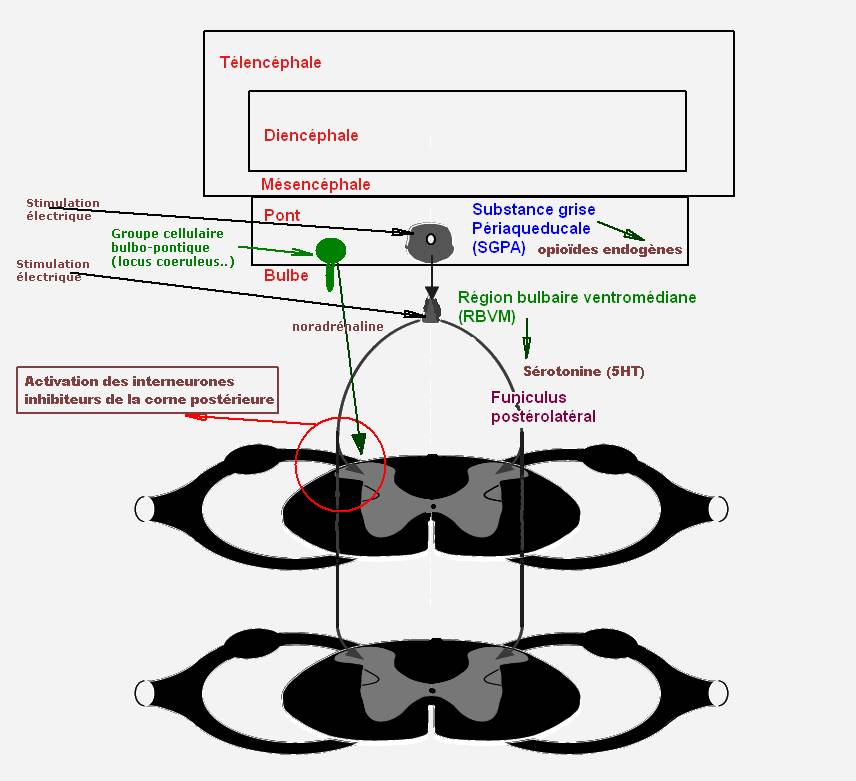
Figure 2. Contrôle inhibiteur descendant faisant intervenir les systèmes à médiation sérotoninergique, opioïdergique et noradrénergique.
Les contrôles facilitateurs descendants supraspinaux
Des contrôles facilitateurs descendants également issus du tronc cérébral, en particulier de la RBVM exacerberaient les conséquences d’une stimulation nociceptive au niveau spinal. Deux types de neurones interviendraient dans la région bulbaire ventromédiane : les neurones « on » excitateurs sur les cellules de transmission (effets facilitateurs proalgiques) et les neurones « off » inhibiteurs antalgiques. L’activation des cellules « off » inhibe la transmission de l’information nociceptive au niveau spinal de la corne postérieure et peut donc être corrélée avec le renforcement de l’inhibition « descendante » de la réponse nociceptive spinale (figure 3) [2,9].
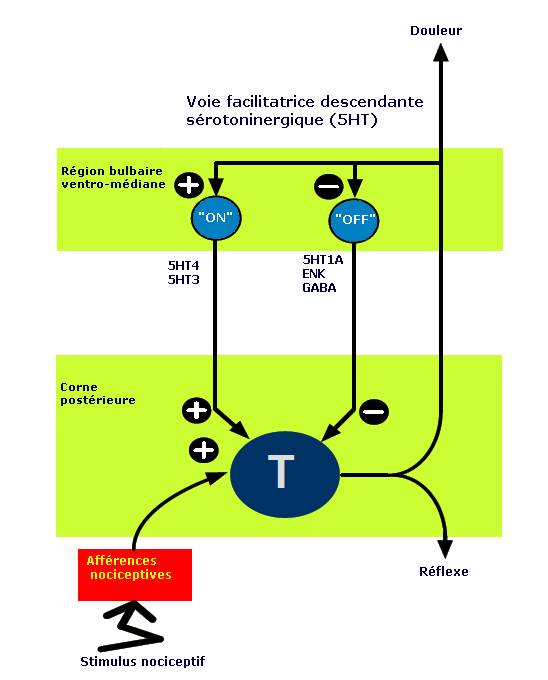
Figure 3. Les contrôles facilitateurs descendants selon Fields.
Les contrôles inhibiteurs diffus induits par une stimulation nociceptive (CIDN)
Si les mécanismes segmentaires antalgiques peuvent bien être engendrés par la stimulation du métamère correspondant, il existe de surcroît un contrôle inhibiteur descendant diffus hypoalgésique engendré par une stimulation nociceptive sur d’autres parties du corps et différentes du site de la douleur initiale. En effet, les neurones à convergence de la corne postérieure sont très fortement inhibés lorsque l’on applique une stimulation nociceptive sur une quelconque partie du corps, différente de leur champ périphérique excitateur : cette stimulation déclenche les CIDN. Les CIDN sont sous-tendus par une boucle complexe faisant intervenir des structures supraspinales appartenant à la formation réticulée bulbaire comme le subnucleus reticularis dorsalis (SRD). Les neuromédiateurs sont endorphiniques et sérotoninergiques (figure 4) [2,8].
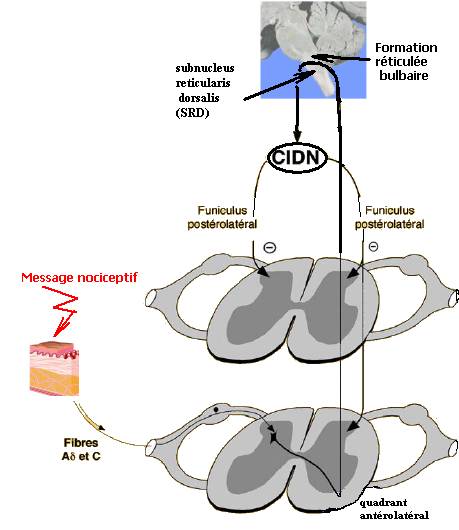
Figure 4. Le CIDN : après stimulation nociceptive quel que soit le métamère, on a une inhibition puissante de tous les segments médullaires (et trigéminaux) non concernés par le stimulus initial (d’après [8]).
Mécanismes d’action de l’acupuncture et de l’électroacupuncture (EA) dans la douleur neuropathique
Les différents opioïdes et leurs récepteurs
En 1992, Chen et Han montrent que l’analgésie produite par l’EA est régulée par trois types de récepteurs opioïdes [10]. Ainsi, l’EA à 2 Hz active les récepteurs μ et δ via les β endorphines, les enképhalines et les endomorphines ; celle à 100 Hz, les récepteurs κ via la dynorphine. L’EA à 15 Hz produit une activation de ces trois sortes de récepteurs chez le rat [11].
On a montré que les effets analgésiques de l’EA sur la douleur nociceptive aiguë sont médiés par les opioïdes endogènes [12], de même que dans les douleurs chroniques [13], l’hyperalgie inflammatoire [14] et les douleurs neuropathiques induites par lésions nerveuses [15].
Cependant il est important de différencier les douleurs nociceptives des douleurs neuropathiques car les effets analgésiques de l’EA varient en fonction de la fréquence.
Il s’avère que les récepteurs opoïdes μ et δ de la corne postérieure de la moelle, stimulés par l’EA à fréquence basse (2Hz) sont impliqués dans l’inhibition de la douleur neuropathique. Par contre, les récepteurs opioïdes κ activés par l’EA à fréquence haute (100Hz) ne le sont pas. Hwang et coll. ont objectivé ainsi sur un modèle de douleur neuropathique chez le rat que la stimulation à 2Hz sur le point houxi (IG3) engendrait un effet antiallodynique réversible par la naloxone [16]. Dans l’allodynie mécanique et l’hyperalgésie thermique induite par lésion nerveuse sur un modèle de rat, on précise que l’EA à 2 Hz a des effets plus importants et plus prolongés que l’EA à 100 Hz [15,17,18].
Par de la capsaïcine injectée à la face plantaire de la patte arrière de rats, Kim et coll. ont créé une hyperalgie persistante par sensibilisation centrale de type neuropathie. Les auteurs démontrent ainsi que l’EA (2Hz, 3mA) appliquée pendant 30mn sur deux points spécifiques (IG3 houxi et TR8 sanyangluo) produit un effet analgésique médié par l’activation des récepteurs opioïdes μ et δ de la corne postérieure de la moelle [19].
L’acupuncture manuelle par stimulation (rotation de l’aiguille à une fréquence de deux tours par seconde pendant 30s) des points sanyinjiao (RA6) et/ou zusanli (ES36) montre son efficacité par inhibition de l’hypersensibilité mécanique sur un modèle animal de douleur neuropathique (ligature des racines lombaires L5-L6 chez le rat). Les auteurs objectivent que le système opioïde est impliqué dans l’efficacité de l’acupuncture manuelle [20].
Une étude expérimentale en utilisant un modèle de rat avec neuropathie périphérique induite par le paclitaxel a évalué l’effet de l’EA sur l’hyperalgésie et l’allodynie mécanique induite par cet agent anticancéreux. L’allodynie mécanique est évaluée par la pression des filaments von Frey (de 4-15 g). L’EA à 10Hz a diminué de manière statistiquement significative (p<0,05) toutes les algies neuropathiques par rapport à l’EA placebo. L’EA à 100Hz va seulement diminuer l’allodynie mécanique à une pression de 15 g. Via les récepteurs opioïdes μ et δ mais non par les récepteurs κ, l’EA à 10Hz inhibe l’allodynie mécanique et l’hyperalgésie paclitaxel-induite plus efficacement que l’EA à 100Hz [21].
Les récepteurs adrénergiques
La noradrénaline (NE) issue du locus coeruleus et des noyaux noradrénergique adjacents au niveau du tronc cérébral sont en rapport avec des récepteurs α2-noradrénergiques et provoque une inhibition directe des neurones de la corne postérieure de la moelle.
L’intervention des récepteurs α2 adrénergiques dans l’analgésie par EA (100Hz) a été observée sur un modèle d’algie nociceptive par entorse chez le rat [22]. De même en 2008, Kim et coll. s’intéressent aux différentes fréquences de stimulation d’EA sur la douleur de type inflammatoire induite par injection de carragénine. Ils observent que l’inflammation est réduite par l’EA à haute ou basse fréquence. L’EA à basse fréquence (1 Hz), tout comme l’acupuncture manuelle avec recherche du deqi (avec rotation à fréquence de 1Hz pendant 90s), est médiée par les ganglions sympathiques post-synaptiques alors qu’à haute fréquence (120 Hz), c’est le système médullaire sympathico-surrénalien qui intervient, sans que la corticostérone ne soit impliquée [23]. Zhang et coll démontrent que, sur un modèle de douleur inflammatoire par injection d’adjuvant de Freund chez le rat, l’analgésie par EA basse fréquence (10Hz) est médiée par les récepteurs α2a adrénergiques [24].
Par contre dans les algies de type neuropathique, l’EA à basse fréquence soulage davantage les douleurs que celle en haute fréquence.
Ainsi, afin d’examiner le rôle des récepteurs α2-adrénergiques dans les mécanismes de l’EA, Kim et al. ont administré une dose d’antagoniste α2-adrénergiques (yohimbine) dans un modèle de douleur neuropathique par section de la racine S1 et S2 innervant la queue du rat. Au préalable, ils avaient stimulé par EA à 2 et à 100Hz le point ES36 (zusanli). Ils montrent que l’EA à 2Hz induit sur l’allodynie au froid un effet analgésique médié par les récepteurs α2-adrénergiques de la moelle plus important que celui médié par l’EA à 100 Hz [25].
En 2007, Li et coll. objectivent que l’EA (10 Hz à 3mA) appliquée pendant 20mn dans un modèle d’algie par injection d’adjuvant complet de Freund (CFA) sur la patte de rat inhibe la transmission nociceptive et hyperalgique en activant les neurones supraspinaux qui se projettent sur la moelle épinière par le système inhibiteur descendant. L’EA active la sérotonine et les catécholamines des neurones du noyau raphé magnus et du locus coeruleus dont l’expression c-fos est significativement augmentée [26].
Les récepteurs cholinergiques
Le système nerveux sympathique fonctionne sur un modèle à deux neurones : le premier pré-ganglionnaire fait synapse avec le second post-ganglionnaire au niveau de la corne postérieure de la moelle. Le neuromédiateur des neurones pré-ganglionnaire est l’acétylcholine alors que celui des neurones post-ganglionnaires est en général la noradrénaline. Il existe deux types de récepteurs cholinergiques : le récepteur nicotinique (récepteur ionotrope perméable aux ions sodium ainsi qu’aux ions K+ sensible à l’acétylcholine) et le récepteur muscarinique (récepteurs métabotropes) tous deux impliqués dans la douleur neuropathique. Cependant, le rôle des récepteurs nicotiniques dans les effets anti-nociceptifs reste plutôt controversé et les récepteurs muscariniques interviendraient davantage [27-29].
Dans la douleur de type inflammatoire, Baek et coll. objectivent ainsi que l’EA à basse fréquence (2Hz, 0,03 ms, 0,07mA) induit un effet analgésique via les récepteurs muscariniques [30].
De la même manière, Park et coll. montrent aussi que les effets antalgiques de l’EA (2Hz ; durée de l’impulsion : 0,5ms) sur l’allodynie au froid et au chaud en rapport avec la douleur neuropathique créée sur un modèle animal par section de racines nerveuse S1 et S2 sont en relation principalement avec les récepteurs muscariniques, en particulier le sous-type M1 [31]. Kim et coll. [[32]] confirment toujours sur le même modèle animal d’algie neuropathique que l’administration intrathécale de néostigmine (parasympathomimétique qui par son effet inhibiteur des cholinestérases, prolonge et augmente les effets nicotiniques et surtout muscariniques de l’acétylcholine) associée à la stimulation d’EA (2 Hz ; 0,5 ms à une intensité de 0,2 -0,3 mA ; 30 mn) produit un effet antalgique synergique qui dure plus de 80 mn.
Les récepteurs sérotoninergiques
L’EA aux fréquences de 2, 10 et 100 Hz a des effets antalgiques dans la douleur de type nociceptive, médiée par les récepteurs 5-HT1A, 5-HT3, mais pas par le récepteur 5-HT2A [33]. Ces observations sont affinées par Baek et coll. qui montrent que l’EA à basse fréquence (2Hz, 0,03 ms, 0,07mA) induit un effet analgésique sur la douleur de type inflammatoire, médié par des récepteurs 5-HT1A et 5-HT3, mais non par le récepteur 5-HT2A chez le rat [30]. Par contre, Takagi et coll. ont déterminé chez le lapin sur un modèle de douleur nociceptive que les récepteurs impliqués dans l’EA à basse fréquence (2Hz) étaient le 5-HT1 (excepté le 5-HT1A), le récepteur 5-HT3, le 5-HT2(excepté le 5-HT2A) [34]. La petite discordance résulte du fait que l’expérimentation a eu lieu dans deux espèces animales différentes, l’une sur un modèle expérimental d’arthrite chez le rat, et l’autre chez le lapin.
Zhang et coll confirment aussi que, sur un modèle de douleur inflammatoire par injection d’adjuvant de Freund chez le rat, l’analgésie par EA basse fréquence (10Hz) est médiée par les récepteurs 5-HT1A au niveau de la corne postérieure de la moelle [24].
Dans la douleur typiquement neuropathique, Kim et coll. suggèrent que l’EA à fréquence basse (2 Hz) est plus appropriée et plus efficace pour le traitement de l’allodynie au froid que l’EA à haute fréquence (100 Hz). Les récepteurs 5-HT1A et 5-HT3 (mais pas le récepteur 5-HT2A)jouent un rôle important dans la médiation des effets antalgiques de la douleur neuropathique à l’allodynie au froid chez le rat [25]. L’EA (10 Hz) active la sérotonine au niveau des neurones du noyau raphé magnus dont l’expression c-fos est significativement augmentée [26].
Les récepteurs GABAergiques
L’un des principaux neuropeptides inhibiteurs, le GABA, libéré dans la substance grise périaqueducale (SGPA) joue un rôle important dans le contrôle inhibiteur descendant de la douleur. Au niveau de la corne postérieure de la moelle épinière, le GABA module la neurotransmission nociceptive entre les neurones afférents primaires et les neurones secondaires des voies spinothalamiques. Les récepteurs GABAA et GABAB contribuent donc principalement à la modulation de la douleur. Ainsi, les agonistes des récepteurs GABAA et GABAB ont des effets anti-nociceptifs [35]. Fusumada et coll ont démontré ainsi que la stimulation par EA (2Hz) au point zusanli (36ES), induisait un effet analgésique chez les rats avec visualisation de l’expression C-Fos au niveau de la SGPA correspondant aux neurones GABAergiques [36].
Dans la douleur de type neuropathique, Park et coll. objectivent sur un modèle animal (section des racines S1 et S2 innervant la queue du rat) que l’EA (2Hz) appliquée au zusanli soulage l’allodynie au froid. Les antagonistes des récepteurs du GABAA et GABAB (respectivement gabazine et saclofène) bloquent l’effet de l’EA à 2Hz signifiant que l’effet antalgique est médié par ces deux récepteurs [37]. Par conséquent, il est possible que le traitement EA à basse fréquence puisse agir en synergie et renforcer les effets analgésiques des médicaments GABAergiques, tels que les agonistes GABA, sur la douleur neuropathique.
Les récepteurs du glutamate (NMDA)
Le glutamate intervient comme neurotransmetteur excitateur qui libéré dans l’espace synaptique, à partir des terminaisons neuronales, se fixe sur ses récepteurs postsynaptiques dont l’activation induit la dépolarisation du neurone cible. Le glutamate agit sur trois récepteurs canaux distincts dénommés par le nom de leur agoniste le plus sélectif : les récepteurs N-méthyl-D-aspartate (NMDA), kainate (KA) et a-amino-3-hydroxy-5-métyl-4-isoxazolepropionate (AMPA). Ces trois récepteurs sont des récepteurs ionotropiques capables de transmettre instantanément un message au neurone cible par modification du potentiel de la membrane post-synaptique en quelques millièmes de secondes.
On a mis en évidence deux familles de récepteurs, respectivement AMPA et kainate (KA) à la fin des années 1970. Les sous-unités AMPA et kainate peuvent être colocalisées au sein d’un même neurone mais ne peuvent s’assembler entre elles. Les différentes sous-unités des récepteurs AMPA/KA sont abondamment exprimées dans l’ensemble du SNC. Les récepteurs au NMDA jouent un rôle important dans différents processus physiologiques en augmentant la transmission du processus douloureux. La kétamine est un antagoniste des récepteurs au NMDA et son injection produit une analgésie puissante.
L’EA (ES36, RA6) à la fréquence de 2 Hz (0,3ms ; 2mA) agit dans les algies nociceptives en diminuant la réponse à l’inflammation locale par l’intermédiaire de la modulation de l’expression des récepteurs ionotropes au glutamate et en particulier le récepteur au NMDA au niveau de la corne dorsale de la moelle épinière [38,39]. Zhang et coll., sur un modèle expérimental animal de douleur inflammatoire par injection de carragénine, montrent que l’EA (2/60Hz) utilisée conjointement avec des antagonistes des récepteurs NMDA ou AMPA/KA possède une action synergétique antinociceptive vis à vis de la douleur inflammatoire [40].
La stimulation des récepteurs α2 adrénergiques par EA à basse fréquence (10Hz) sur un modèle de douleur inflammatoire par injection de CFA chez le rat diminue la libération de glutamate au niveau de la corne postérieure de la moelle par régulation de la sous-unité NR1 des récepteurs NMDA. De ce fait, la NE va engendrer une diminution de la libération pré-synaptique de glutamate, conduisant ainsi à l’inhibition de la douleur de type inflammatoire [24].
Wang et coll. ont confirmé en 2006 le rôle de la modulation de l’expression des récepteurs de la NMDA par EA (4 Hz/16 Hz) sur un modèle de rat hyperalgique induit par injection de CFA, jouant un rôle inhibiteur du développement de la sensibilisation centrale en rapport avec la douleur inflammatoire chronique [41].
Concernant la douleur neuropathique proprement dite, Xing et coll. a démontré que l’EA à basse fréquence (2 Hz) intervient dans la plasticité synaptique rachidienne par une dépression à long terme de la transmission synaptique au niveau de la corne dorsale de la moelle épinière contribuant aux effets analgésiques de longue durée. Ces effets de l’antinociception s’expliqueraient par l’induction des récepteurs NMDA via l’activation du système des peptides opioïdes endogènes [42].
Conclusion
L’analgésie induite par acupuncture ou électroacupuncture de la douleur neuropathique fait intervenir, comme dans la douleur nociceptive [12], différents mécanismes entraînant une modulation de la sensibilisation périphérique, centrale et de la plasticité synaptique au niveau du SNC [1] via les contrôles inhibiteurs spinaux et supraspinaux. Le rôle des systèmes opioïdes (récepteurs opioïdes μ et δ), adrénergiques (récepteur α2-adrénergique), sérotoninergiques (récepteurs 5-HT1A et 5-HT3), cholinergiques (récepteur muscarinique M1) et GABAergiques (récepteurs GABAA et GABAB) est prépondérant [43] (figure 5). L’EA à basse fréquence est plus efficace que celle à haute fréquence dans les douleurs neuropathiques.
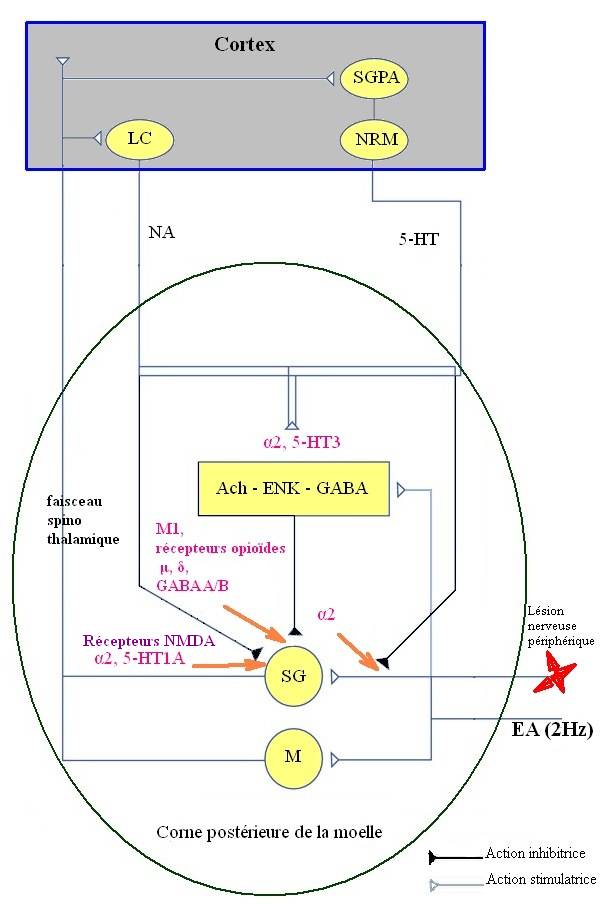
Figure 5. Modulation et contrôle de la douleur neuropathique par l’EA au niveau du SNC. La douleur neuropathique est transmise par les fibres afférentes primaires via la substance gélatineuse (SG). L’EA engendre une stimulation par les cellules (M) marginales correspondant aux neurones T de la théorie du « portillon » via le faisceau spino-thalamique, où le signal est transmis vers le cortex. Il y a implication des neurones cholinergiques, enképhalinergiques et GABAergiques. La SGPA projette des neurones vers le NRM qui envoie à son tour des fibres sérotoninergiques (5-HT) vers la corne dorsale. Le LC envoie également des fibres noradrénergiques vers la corne dorsale. La NE, par l’intermédiaire des récepteurs α2 -adrénergiques accentue l’action des neurones cholinergiques et GABAergiques de la moelle et engendre une diminution de la libération de transmetteurs pronociceptifs au niveau des fibres afférentes primaires. Elle inhibe aussi la transmission des signaux de la douleur au niveau des afférences secondaires supraspinales. La sérotonine active les enképhalines (ENK) et le GABA des neurones intrinsèques à travers les récepteurs sérotoninergiques à 5-HT3 et inhibe les afférences secondaires par les récepteurs sérotoninergiques 5-HT1A. Les neurones cholinergiques, enképhalinergiques et GABAergiques, par le biais des récepteurs muscariniques M1, des récepteurs opioïdes μ, δ, des récepteurs NMDA et des récepteurs GABAA et GABAB contrôlent les entrées nociceptives de la périphérie vers le cortex. SG : substance gélatineuse ; M : cellules marginales ; SGPA : substance grise périaqueducale ; NRM : noyau du raphé magnus ; LC: locus coeruleus ; 5-HT : la sérotonine ; NE: noradrénaline ; Ach : acétylcholine ; ENK : enképhaline. Schéma d’après [43].
Références
[1]. Stéphan JM. Neuropathies périphériques induites par la chimiothérapie : mécanismes d’action de l’acupuncture dans la sensibilisation périphérique et centrale. Acupuncture & Moxibustion. 2013;12(4):305-314.
[2]. Calvino B, Grilo RM. Central pain control. Joint Bone Spine. 2006;73(1):10-6.
[3].Garcia-Larrea L, Magnin M. [Pathophysiology of neuropathic pain: review of experimental models and proposed mechanisms]. Presse Med. 2008;37(2 Pt 2):315-40.
[4]. Zhuo M. Neuronal mechanism for neuropathic pain. Mol Pain. 2007 Jun 6;3:14.
[5]. Melzack R,Wall PD. Pain mechanisms: a new theory. Sci. 1965;150:971–9.
[6]. Wall PD. The gate control theory of pain mechanisms: a re-examination and re-statement. Brain. 1978;101:1-18.
[7]. Fields HL. Pain modulation: expectation, opioid analgesia and virtual pain. Prog Brain Res 2000;122:245-53.
[8]. Le Bars D, Willer JC. Physiologie de la douleur. EMC – Anesthésie-Réanimation. 2004;1(4):227-266.
[9]. Fields HL. Is there a facilitating component to central pain modulation? Am Pain Soc J 1992;1:71–8.
[10]. Chen XH, Han JS. Analgesia induced by electroacupuncture of different frequencies is mediated by different types of opioid receptors: another cross-tolerance study. Behav Brain Res. 1992;47(2):143-9.
[11] . Han JS. Acupuncture and endorphins. Neurosci Lett. 2004;361(1-3):258-61.
[12]. Stéphan JM. Mécanismes neurophysiologiques de l’électroacupuncture dans les algies. Acupuncture & Moxibustion. 2008;7(2):127-137.
[13]. Lao L, Zhang RX, Zhang G, Wang X, Berman BM, Ren K. A parametric study of electroacupuncture on persistent hyperalgesia and Fos protein expression in rats. Brain Res. 2004;1020(1-2):18-29.
[14]. Yang EJ, Koo ST, Kim YS, Lee JE, Hwang HS, Lee MS, Choi SM. Contralateral electroacupuncture pretreatment suppresses carrageenan-induced inflammatory pain via the opioid-mu receptor. Rheumatol Int. 2011 Jun;31(6):725-30.
[15]. Kim JH, Min BI, Na HS, Park DS. Relieving effects of electroacupuncture on mechanical allodynia in neuropathic pain model of inferior caudal trunk injury in rat: mediation by spinal opioid receptors. Brain Res. 2004 Feb 20;998(2):230-6.
[16]. Hwang BG, Min BI, Kim JH, Na HS, Park DS. Effects of electroacupuncture on the mechanical allodynia in the rat model of neuropathic pain. Neurosci Lett. 2002;320(1-2):49-52.
[17]. Sun RQ, Wang HC, Wang Y. [Effect of electroacupuncture with different frequencies on neuropathic pain in a rat model]. Zhongguo Ying Yong Sheng Li Xue Za Zhi. 2002 May;18(2):128-31.
[18]. Han JS. Acupuncture: neuropeptide release produced by electrical stimulation of different frequencies. Trends Neurosci. 2003 Jan;26(1):17-22
[19]. Kim HY, Wang J, Lee I, Kim HK, Chung K, Chung JM. Electroacupuncture suppresses capsaicin-induced secondary hyperalgesia through an endogenous spinal opioid mechanism. Pain. 2009 Oct;145(3):332-40.
[20]. Cidral-Filho FJ, da Silva MD, Moré AO, Córdova MM, Werner MF, Santos AR. Manual acupuncture inhibits mechanical hypersensitivity induced by spinal nerve ligation in rats. Neuroscience. 2011;193:370-6.
[21]. Meng X, Zhang Y, Li A, Xin J, Lao L, Ren K, Berman BM, Tan M, Zhang RX. The effects of opioid receptor antagonists on electroacupuncture-produced anti-allodynia/hyperalgesia in rats with paclitaxel-evoked peripheral neuropathy. Brain Res. 2011;1414:58-65.
[22]. Koo ST, Lim KS, Chung K, Ju H, Chung JM. Electroacupuncture-induced analgesia in a rat model of ankle sprain pain is mediated by spinal alpha-adrenoceptors. Pain. 2008;135(1-2):11-9.
[23]. Kim HW, Uh DK, Yoon SY, Roh DH, Kwon YB, Han HJ, Lee HJ, Beitz AJ, Lee JH. Low-frequency electroacupuncture suppresses carrageenan-induced paw inflammation in mice via sympathetic post-ganglionic neurons, while high-frequency EA suppression is mediated by the sympathoadrenal medullary axis. Brain Res Bull. 2008 Mar 28;75(5):698-705.
[24]. Zhang Y, Zhang RX, Zhang M, Shen XY, Li A, Xin J, Ren K, Berman BM, Tan M, Lao L. Electroacupuncture inhibition of hyperalgesia in an inflammatory pain rat model: involvement of distinct spinal serotonin and norepinephrine receptor subtypes. Br J Anaesth. 2012 Aug;109(2):245-52.
[25]. Kim SK, Park JH, Bae SJ, Kim JH, Hwang BG, Min BI, Park DS, Na HS. Effects of electroacupuncture on cold allodynia in a rat model of neuropathic pain: mediation by spinal adrenergic and serotonergic receptors. Exp Neurol. 2005;195(2):430-6.
[26]. Li A, Wang Y, Xin J, Lao L, Ren K, Berman BM, Zhang RX. Electroacupuncture suppresses hyperalgesia and spinal Fos expression by activating the descending inhibitory system. Brain Res. 2007 Dec;1186:171-9.
[27]. Umana IC, Daniele CA, McGehee DS. Neuronal nicotinic receptors as analgesic targets: It’s a winding road. Biochem Pharmacol. 2013;86(8):1208-14.
[28]. Cai YQ, Chen SR, Han HD, Sood AK, Lopez-Berestein G, Pan HL. Role of M2, M3, and M4 muscarinic receptor subtypes in the spinal cholinergic control of nociception revealed using siRNA in rats. J Neurochem. 2009;111(4):1000-10.
[29]. Obata H, Saito S, Sasaki M, Goto F. Possible involvement of a muscarinic receptor in the anti-allodynic action of a 5-HT2 receptor agonist in rats with nerve ligation injury. Brain Res. 2002;932(1-2):124-8.
[30]. Baek YH, Choi DY, Yang HI, Park DS. Analgesic effect of electroacupuncture on inflammatory pain in the rat model of collagen-induced arthritis: mediation by cholinergic and serotonergic receptors. Brain Res. 2005 Sep 28;1057(1-2):181-5.
[31]. Park JH, Kim SK, Kim HN, Sun B, Koo S, Choi SM, Bae H, Min BI. Spinal cholinergic mechanism of the relieving effects of electroacupuncture on cold and warm allodynia in a rat model of neuropathic pain. J Physiol Sci. 2009;59(4):291-8.
[32]. Kim HN, Park JH, Kim SK, Sun B, Koo S, Choi SM, Bae H, Min BI. Electroacupuncture potentiates the antiallodynic effect of intrathecal neostigmine in a rat model of neuropathic pain. J Physiol Sci. 2008;58(5):357-60.
[33]. Chang FC, Tsai HY, Yu MC, Yi PL, Lin JG. The central serotonergic system mediates the analgesic effect of electroacupuncture on zusanli (ST36) acupoints. J Biomed Sci. 2004;11(2):179-85.
[34]. Takagi J, Yonehara N. Serotonin receptor subtypes involved in modulation of electrical acupuncture. Jpn J Pharmacol. 1998;78(4):511-4.
[35]. Bardoni R, Takazawa T, Tong CK, Choudhury P, Scherrer G, Macdermott AB. Pre-and postsynaptic inhibitory control in the spinal cord dorsal horn. Ann N Y Acad Sci. 2013 Mar;1279:90-6.
[36]. Fusumada K, Yokoyama T, Miki T, Wang ZY, Yang W, Lee NS, Endo Y, Takeuchi Y. c-Fos expression in the periaqueductal gray is induced by electroacupuncture in the rat, with possible reference to GABAergic neurons. Okajimas Folia Anat Jpn. 2007;84(1):1-9.
[37]. Park JH, Han JB, Kim SK, Park JH, Go DH, Sun B, Min BI. Spinal GABA receptors mediate the suppressive effect of electroacupuncture on cold allodynia in rats. Brain Res. 2010;1322:24-9.
[38]. Choi BT, Lee JH, Wan Y, Han JS. Involvement of ionotropic glutamate receptors in low frequency electroacupuncture analgesia in rats. Neurosci Lett. 2005;377(3):185-8.
[39]. Choi BT, Kang J, Jo UB. Effects of electroacupuncture with different frequencies on spinal ionotropic glutamate receptor expression in complete Freund’s adjuvant-injected rat. Acta Histochem. 2005;107(1):67-76.
[40]. Zhang YQ, Ji GC, Wu GC, Zhao ZQ. Excitatory amino acid receptor antagonists and electroacupuncture synergetically inhibit carrageenan-induced behavioral hyperalgesia and spinal fos expression in rats. Pain. 2002 Oct;99(3):525-35.
[41]. Wang L, Zhang Y, Dai J, Yang J, Gang S. Electroacupuncture (EA) modulates the expression of NMDA receptors in primary sensory neurons in relation to hyperalgesia in rats. Brain Res. 2006;1120(1):46-53.
[42]. Xing GG, Liu FY, Qu XX, Han JS, Wan Y. Long-term synaptic plasticity in the spinal dorsal horn and its modulation by electroacupuncture in rats with neuropathic pain. Exp Neurol. 2007;208(2):323-32.
[43]. Kim W, Kim SK, Min BI. Mechanisms of electroacupuncture-induced analgesia on neuropathic pain in animal model. Evid Based Complement Alternat Med. 2013;2013:436913.