
Résumé : Introduction. L’objectif de ce travail est d’évaluer la possibilité d’utiliser l’électroacupuncture (EA) chez la femme enceinte dans les nausées-vomissements, la maturation cervicale et l’induction du travail. Méthodes. A partir de deux cas cliniques, la discussion portera sur l’intérêt d’utiliser l’EA en obstétrique en complément de l’acupuncture manuelle en se basant sur un état des lieux des essais comparatifs randomisés (ECR) et des méta-analyses. L’acupuncture expérimentale est abordée également, permettant de mieux appréhender les mécanismes neurophysiologiques. Résultats. L’utilisation de l’EA ou de la neurostimulation électrique transcutanée appliquée aux points d’acupuncture (TEAS) potentialise les effets de l’acupuncture manuelle seule. Conclusion. Selon les preuves issues des ECR, l’EA peut être considérée comme plus efficace dans les nausées et vomissements, la maturation du col utérin et l’induction du travail chez la femme enceinte que l’acupuncture manuelle isolée. Mots clés : Electroacupuncture – Nausées – Vomissements – Maturation du col – Induction du travail – Obstétrique – Neurostimulation électrique transcutanée appliquée aux points d’acupuncture – TEAS – Mécanismes neurophysiologiques.
Summary: Introduction. The objective of this study was to evaluate the possibility of using electroacupuncture (EA) in pregnant women in nausea, vomiting, cervical ripening and induction of labor. Methods. From two clinical cases, discussion will focus on the benefits of using EA in obstetrics in addition to manual acupuncture based on an inventory of randomized controlled trials (RCTs) and meta-analyzes. The experimental acupuncture is also addressed, to better understand the neurophysiological mechanisms. Results. Using the EA or transcutaneous electrical acupoint stimulation (TEAS) potentiates the effects of manual acupuncture alone. Conclusion. According to evidence from RCTs, the EA can be considered more effective in nausea and vomiting, ripening the cervix and inducing labor in pregnant women that the only manual acupuncture. Keywords: Electroacupuncture – Nausea – Vomiting – Maturation of the cervix – Induction of labor – Obstetrics – Transcutaneous Electrical Nerve Stimulation applied to acupuncture points – TEAS – neurophysiological mechanisms.

Nous avons vu dans un précédent article la place de l’électroacupuncture (EA) [[1]] dans les algies chez la femme enceinte. Nous abordons à partir de cas cliniques, son intérêt dans les pathologies de la grossesse en dehors de l’action analgésique avec les modalités des protocoles confrontés aux données issues des essais comparatifs randomisés (ECR).
Cas cliniques
Nausées et vomissements du premier trimestre
Une jeune femme de 28 ans, primipare, consulte à 15 semaines d’aménorrhée (SA) pour des nausées opiniâtres accompagnées d’hypersialorrhée, de crachats mousseux et de vomissements clairs de glaires et salive. La langue est épaisse avec empreintes des dents et enduit lingual central blanc, gras. Le pouls est glissant, perlé (hua). Selon la différenciation des syndromes (bianzheng), un diagnostic de Mucosités-Glaires est porté. Trois séances de 20mn à une semaine d’intervalle entraînent la disparition complète des nausées et vomissements. Les points utilisés : 6MC (neiguan), 12VC (zhongwan), 40E (fenglong) stimulé par EA à une fréquence de 100Hz (300µs), 3Rt (taibai) et 36E (zusanli).
Maturation du col et induction du travail
Chez cette seconde pare à 41SA, avec un petit retard d’un jour par rapport à la date présumée d’accouchement et sans contractions utérines, une induction du travail est souhaitée. Un traitement non individualisé standard est utilisé impliquant les points 3F (taichong), 4GI (hegu), 6Rt (sanyinjiao), 31V (shangliao) et 32V (ciliao), 3VC (zhongji) et 4VC (guanyuan), les quatre derniers points en EA à une fréquence de 2Hz (300µs) pendant 30 minutes. Quelques contractions surviennent durant la séance et l’accouchement survient 20h après.
L’électroacupuncture à visée antiémétique
Le seul et unique ECR concernant l’EA dans les nausées et vomissements (NV) de la femme enceinte et qui démontre son efficacité est celui de Rosen [[2]]. Cet ECR en simple aveugle de bonne qualité méthodologique (n=187) montre une réduction au cours du premier trimestre de la grossesse de l’EA (fréquence non connue) sur 6MC (neiguan) versus groupe placebo. En fait, il s’agirait davantage d’un dispositif de neurostimulation électrique transcutanée appliquée aux points d’acupuncture (TEAS = transcutaneous electrical acupoint stimulation) car la stimulation électrique se ferait par l’intermédiaire d’un bracelet Velcro de 34g muni d’électrodes adhésives appliquées sur la peau. La TEAS est réalisée par l’intermédiaire d’électrodes adhésives appliquées sur la peau de manière transcutanée alors que l’EA est percutanée. Quoiqu’il en soit et selon la collaboration Cochrane de Matthews et coll. [[3]], la stimulation électrique serait plus efficace que l’acupuncture manuelle.
Pour se faire une idée de l’efficacité de l’EA dans les NV, il faut se rapprocher aussi de ce qui se fait dans les NV induits par la chimiothérapie. Ainsi, la méta-analyse de Ezzo et coll. qui a étudié les trois différentes formes d’interventions (acupuncture manuelle, EA et acupression) objective que l’EA dans trois ECR (n=134) est bénéfique dans les vomissements aigus versus le traitement habituel (p=0,02). Versus groupe témoin, l’acupuncture manuelle ne serait pas efficace dans les NV et l’acupression le serait dans les nausées aiguës mais pas dans les vomissements [[4]]. L’EA est utilisée aux points 36E (zusanli) et 6MC (neiguan) à une fréquence de 2 à 10Hz (impulsion de 500µs à 700µs) [[5]].
Acupuncture expérimentale
Expérimentalement chez la souris, on observe que l’EA (2Hz alternée à 15Hz) au point 12VC (zhongwan) réduit la motilité gastrique par action sur les récepteurs vanilloïdes 1 (TRPV1) [[6]]. Une autre théorie suggère que chez le chien, l’EA au 6MC (1 à 30Hz) réduit les contractions rétrogrades péristaltiques à travers l’action directe sur les muscles lisses de l’intestin grêle et de l’estomac, action abolie par la naloxone, d’où l’implication des neuropeptides opioïdes [[7]]. Idem chez l’homme avec une EA à 1Hz, mais avec une modulation de l’activité myoélectrique gastrique visible par électro-gastrographie (EGG). Ainsi, l’EA sur le 6MC inhibe l’amplitude du péristaltisme gastrique alors que le 36E l’amplifie. Par contre, s’ils sont stimulés ensemble, on a un effet synergétique par diminution du péristaltisme gastrique [[8]].
Chez quatorze sujets sains dont on a déclenché volontairement une distension gastrique, la stimulation électrique du 6MC par TEAS (100Hz avec impulsion de 100µs) réduit de manière significative (40% ; p<0,02) la relaxation transitoire du sphincter inférieur de l’œsophage (RTSIO)[a], action non inhibée par la naloxone chez l’homme. Cet effet ne semble pas être médié par les récepteurs µ opioïdes [[9]]. On retrouve le même résultat chez le chat. L’EA à une fréquence de 2Hz alternant avec celle de 100Hz (2/100Hz) sur le 6MC et 36E réduit la RTSIO et peut être médiée par l’oxyde nitrique (NO)[b], les récepteurs CCK-A et les récepteurs µ opioïdes [[10]] et les voies cholinergiques [[11],[12]].
Commentaires sur le cas clinique
Selon les conceptions de la Médecine Chinoise, la croissance progressive du fœtus enclenche une augmentation progressive du qi, lequel associé à l’accumulation du qi du chongmai va déclencher une remontée de qi à contre-courant. Ce mécanisme contrarie le mouvement de descente normale du qi de l’Estomac qui va alors s’épuiser progressivement. Dans ce cas clinique, il y a un Vide de qi de la Rate et l’Humidité s’accumule et entraîne au final la formation de Mucosités-Glaires (tanyin). Le point important à puncturer est donc le 40E. Selon les données expérimentales réalisées chez l’homme, mais aussi chez l’animal, l’EA à une fréquence de 100Hz ou 2/100Hz semble le meilleur compromis pour réduire la RTSIO. En cas d’un bianzheng évoquant un Vide de qi de Cœur et Feu du Cœur, le 6MC en EA à une fréquence de 2Hz aurait été plus judicieux.
L’électroacupuncture dans la maturation du col puis l’induction du travail
La provocation des contractions utérines ne suffit pas pour déclencher l’accouchement. On sait que pour se dilater, un col de l’utérus doit subir des modifications de structure. De ce fait, la méthode de déclenchement dépendra de l’état du col de l’utérus à apprécier par le toucher vaginal grâce au score probablement le plus utilisé en obstétrique, le score de Bishop (figure 1).
| Évaluation de la maturation du col utérin selon le score de Bishop (1964)[c] | ||||
| 0 | 1 | 2 | 3 | |
| Dilatation du col | fermé | 1-2 cm | 3-4 cm | ≥ 5 cm |
| Position | Postérieure | Centrale | Antérieure | |
| Effacement | Long (0-30%) | Mi-long (40-50%) | Court (60-70%) | Effacé (>80%) |
| Consistance | Ferme | Moyenne | Molle | |
| Présentation (Positionnement de la présentation fœtale par rapport aux épines sciatiques) | Mobile (3 cm au dessus) | Amorcée (2 cm au dessus) | Fixée (<1cm au dessus) | Engagée |
Figure 1. Valeurs du score : de 0 à 13 ; score >= à 7 : pronostic favorable (travail de moins de 4 heures chez les multipares).
Lorsque le score de Bishop est favorable, le déclenchement du travail se fait directement. En cas de score défavorable, il est alors nécessaire tout d’abord de réaliser une maturation du col qui sera ensuite suivi du déclenchement.
La méta-analyse Cochrane de Smith et coll.
La Haute Autorité de Santé préconise la maturation cervicale par l’utilisation des prostaglandines E2 sous forme intravaginale, préférable à celle de l’ocytocine, du misoprostol (prostaglandine E1) et de la mifépristone pour le déclenchement artificiel du travail. En 2008, la HAS notifiait que les données disponibles ne permettaient pas de conclure sur l’intérêt de l’utilisation de l’acupuncture pour induire le travail[d]. Elle se basait sur la revue Cochrane de 2004 [[13]] et deux essais comparatifs randomisés [[14],[15]] qui avait pour objectif d’évaluer les effets de l’acupuncture sur la maturation cervicale et le déclenchement du travail pendant le troisième trimestre de grossesse. La revue Cochrane réalisée en 2004 et réactualisée en 2008 concluait qu’on avait besoin d’essais contrôlés randomisés bien conçus et de haute qualité méthodologique pour évaluer le rôle de l’acupuncture dans l’induction du travail avec des résultats cliniquement significatifs. Pourtant, les auteurs s’appuyaient sur trois ECR (n=212) [14,15,[16]]. Smith et al. objectivaient une efficacité clinique de l’acupuncture statistiquement significative (147 femmes, risque relatif RR=1,45 avec un intervalle de confiance IC à 95% de 1,08- 1,95 ; p=0,01) par rapport au groupe contrôle relatant que l’acupuncture nécessitait moins l’utilisation des autres techniques d’induction versus les groupes contrôles. Néanmoins, les auteurs concluaient que la population incluse était trop petite et que les femmes n’étaient pas décrites comme aveugles dans leur groupe. De ce fait, les résultats pouvaient être dus à un effet placebo [[17]].
Cette revue bénéficia d’une mise à jour en 2013 [[18]] avec inclusion de onze nouveaux ECR, soit quatorze ECR (n=2220 femmes). Trois critères de jugement principaux ont été étudiés : césarienne, morbidité néonatale grave et mortalité maternelle. Aucun essai ne documentait d’accouchement non conclu par voie basse dans les 24 heures et d’hyperstimulation utérine avec des modifications de la fréquence cardiaque fœtale (FCF).
On objectivait qu’il y avait certaines preuves d’un changement dans la maturation cervicale pour les femmes recevant de l’acupuncture par rapport au groupe témoin sous acupuncture simulée[e] (avec une différence moyenne (DM) : 0,40 ; IC à 95% = 0,11-0,69 ; p=0,0062 dans un ECR de 125 femmes) [[19]] et versus soins habituels avec misoprostol (DM : 1,30 ; IC 95% = 0,11-2,49 ; p=0,032) dans un ECR de 67 femmes [[20]] (figure 2). Remarquons que les auteurs de la Cochrane signalaient qu’une étude (Trémeau 1992 [[21]]) avait également apporté de plus grands changements dans la maturation du col de l’utérus dans le groupe acupuncture par rapport aux soins habituels, mais ne les avaient pas inclus dans la méta-analyse, du fait de l’hétérogénéité des protocoles.
Par contre, il n’y avait aucune autre différence statistiquement significative dans six ECR (n=654) en termes d’accouchements par césarienne entre groupe acupuncture et groupe témoin sous acupuncture simulée (risque relatif moyen (RR) : 0,95 ; IC à 95 % = 0,69 à 1,30 ; p=0,65) [19,21,[22],[23],[24],26]. Pas de différence non plus entre groupe acupuncture et groupe soins habituels dans six ECR (n=364) (RR moyen : 0,69 ; IC à 95 % = 0,40-1,20 ; p=0,19) [15,20,21,22,24,[25]] pour les accouchements par césarienne. Dans un ECR de 67 femmes [20], la durée du travail était plus courte dans le groupe de soins habituels (misoprostol) par rapport à l’acupuncture (moyenne de la différence moyenne standardisée DMS = 0,67 ; IC 95% = 0,18-1,17, p=0,0076).
A noter qu’il n’y avait pas de différence statistiquement significative dans les convulsions néonatales entre groupe acupuncture et groupe sous acupuncture simulée dans un essai (n=364) [[26]] (RR : 1,01 ; IC à 95% = 0,06 – 16,04 ; p=0,99), objectivant la sécurité de l’acupuncture.
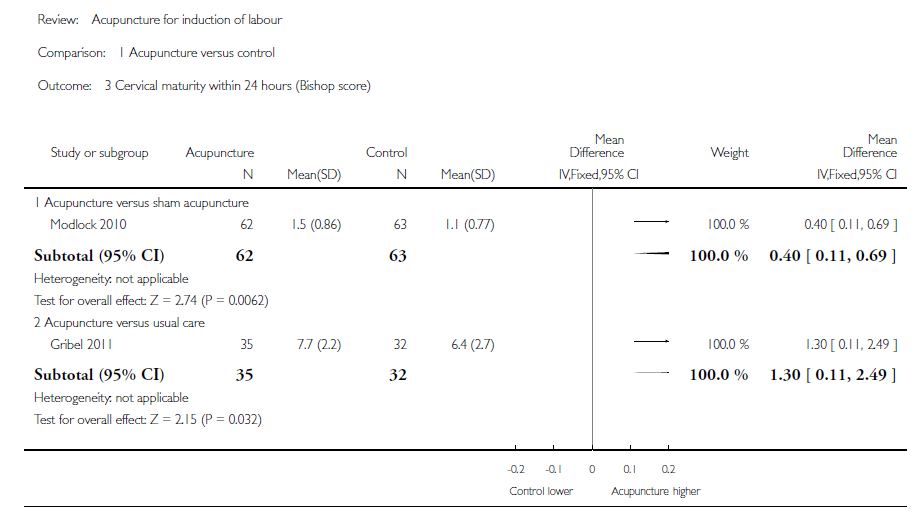
Figure 2. Comparaison de l’acupuncture versus groupe témoin dans la maturité du col utérin dans les 24h par évaluation du score de Bishop [18].
Depuis cette méta-analyse Cochrane de 2013, trois ECR sont parus [27-29]. Ajori et coll. (n=80 femmes à 38SA) utilisant le protocole d’acupuncture (4GI, 6Rt, 67V) n’ont pas objectivé de maturation ou d’induction du travail. Même chose avec l’ECR d’Andersen et coll. (n=407 femmes à 41SA)[f], et celui de Neri et coll. (n=221 femmes à 41SA + 5 jours) qui ont planifié des séances d’acupuncture tous les deux jours pendant une semaine.
Electroacupuncture
Comme le laisse entendre la méta-analyse Cochrane, la stimulation électrique des points d’acupuncture (EA) offre une maturation et une induction du travail plus efficace que l’acupuncture manuelle.
Ainsi, dans trois études chinoises, la majorité des femmes enceintes en terme dépassé avaient commencé le travail pendant le traitement électroacupunctural (fréquences entre 2 et 8Hz sur les points 6Rt et 4GI). Cependant, aucune de ces études n’avait inclus de groupe témoin, d’où le risque majeur de biais [17]. On notera aussi que la neurostimulation électrique transcutanée appliquée aux points d’acupuncture (TEAS = transcutaneous electrical acupoint stimulation) augmente la fréquence et la force des contractions utérines [[30]] (figure 3).
En 2006, dans l’ECR de Harper et coll. [15] l’EA à 2Hz montre une tendance à un accouchement plus rapide dans le groupe acupuncture par rapport au groupe témoin (p=0,36) ainsi qu’une tendance à avoir moins de césariennes. Mais il existe des limitations de ce travail liées à l’inclusion d’un petit nombre de femmes (manque de puissance) et une étude réalisée non en insu (figure 4).

Figure 3. Le protocole de TEAS de Dunn et coll. [30] utilisant la stimulation électrique à la fréquence de 30Hz.
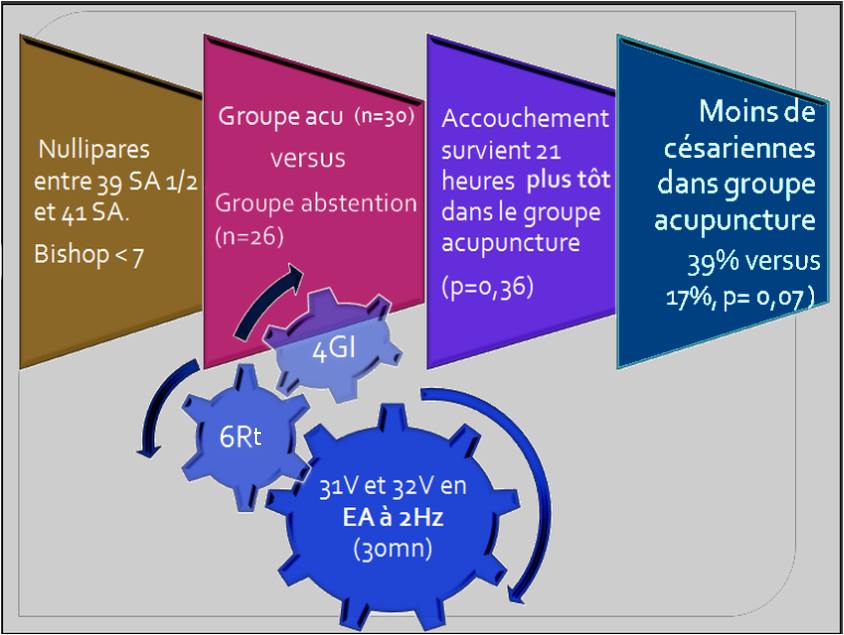
Figure 4. La punture des points 4GI (hegu), 6Rt (sanyinjiao) et l’EA (2Hz) appliquée aux 31V (shangliao) et 32V (ciliao) entraine un accouchement survenu 21 heures plus tôt dans le groupe acupuncture par rapport au groupe contrôle, mais statistiquement non significatif (p=0,36) et moins de césariennes que le groupe témoin [15].
Un autre ECR canadien [23] de Gaudet et coll. en 2008 a essayé de déterminer l’efficacité de l’acupuncture pour induire le travail chez les femmes à terme à 41 SA (282 jours de grossesse en moyenne). Les auteurs ont constaté une différence de 62 heures, pour ce qui est de l’intervalle séparant l’intervention et l’accouchement, entre les deux groupes (en faveur du groupe « traitement ») et des périodes de travail plus courtes (figure 5). Cette étude pilote en double insu contre placebo de haute qualité méthodologique est malheureusement de très faible puissance. Les auteurs ont d’ailleurs calculé que pour avoir une puissance à 80% avec un risque alpha α de 5%, il fallait trente-huit parturientes par groupe pour détecter une différence.
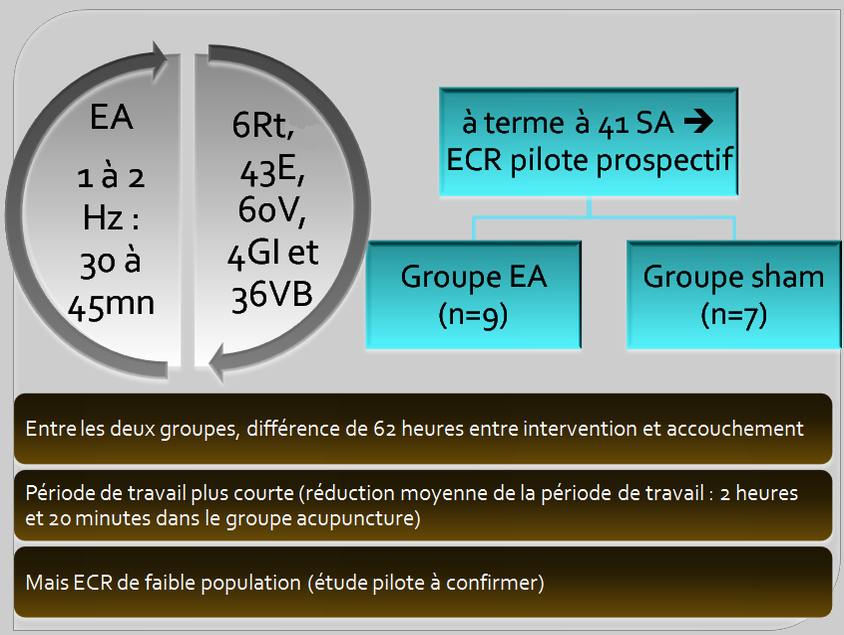
Figure 5. L’EA à la fréquence variant de 1 à 2Hz est appliquée sur 6Rt, 43E (xiangu), 60V, 4GI et 36VB (waiqiu). Le groupe sham (acupuncture simulée) reçoit l’acupuncture sur des sites adjacents mais en dehors des méridiens et stimulés également par EA [23].
L’ECR de Gribel et coll. [20] paru en 2011 au Brésil n’est pas en insu tant pour les patientes que les praticiens et vise à comparer les effets de l’utilisation de l’EA (5/50Hz) et du misoprostol dans l’induction du travail chez les parturientes ayant un score de Bishop inférieur à 7.
Il n’y a pas de différence significative du travail dans les deux groupes en ce qui concerne la fréquence (p = 0,07) et le temps d’induction (p = 0,29), ce qui objective que l’EA est aussi efficace que le misoprostol. Cependant, on note surtout une absence de complication obstétricale et une plus grande satisfaction des patientes (p = 0,046) observées chez les patients du groupe EA bien que la durée du travail (p = 0,036) soit plus longue. Il existe aussi une fréquence plus élevée de césariennes (p = 0,014) et les complications obstétricales (9,3%) ont été observés chez les patientes du groupe misoprostol.
En conclusion, on objective que l’EA offre des résultats similaires dans la maturation du col et l’induction du travail au misoprostol mais sans survenue de complications obstétricales (figure 6).
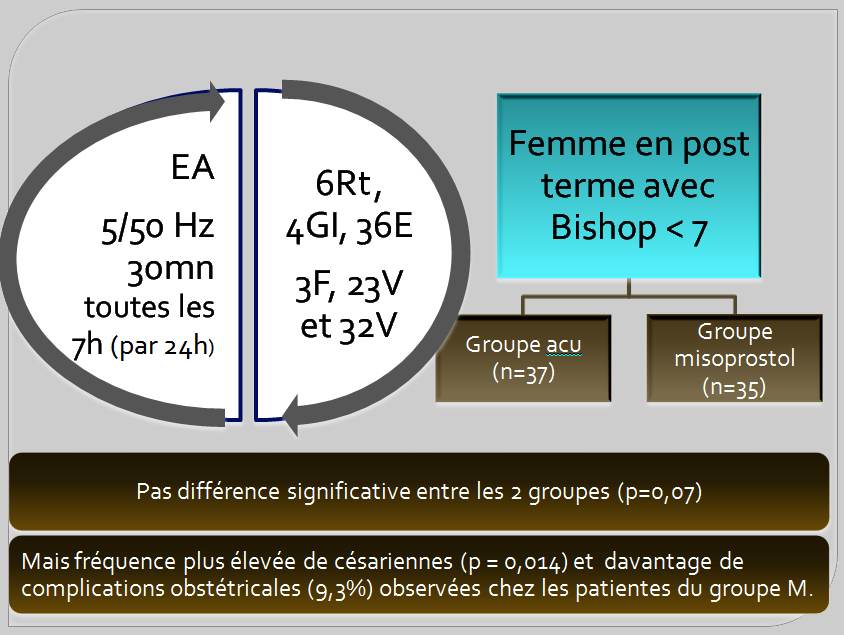
Figure 6. L’EA (alternance 5/50Hz ; durée d’impulsion 200µs) aux points hegu (4GI), sanyinjiao (6Rt), zusanli (36E), taichong (3F), shenshu (23V) et ciliao (32V) peut être utilisée pour obtenir la maturation du col, avec des résultats similaires au misoprostol, avec une fréquence significativement plus élevée d’accouchements par voie basse et sans survenue de complications obstétricales [20]. L’EA a été exécutée tous les 7 h durant une hospitalisation par période de 24 h par cure de 3 sessions.
Paramètres électrophysiologiques
Une étude d’électroacupuncture expérimentale chez des rates au dernier stade de la grossesse a permis de déterminer les paramètres d’efficacité pour induire le travail en mesurant les contractions utérines. Les auteurs ont objectivé que l’EA sur hegu (4GI) et sanyinjiao (6Rt) pendant 20mn, entraîne des contractions utérines statistiquement significatives dans tous les groupes d’EA (n= 12 dans chaque groupe ; groupe EA traité par EA à 15Hz, groupe EA fréquence 2Hz en alternance 50 Hz, groupe 30Hz, groupe 50Hz, groupe 2/15Hz et 2/30Hz) par rapport au groupe témoin (animaux sans grossesse). On constate néanmoins que le groupe EA 2/50Hz offre une moyenne des niveaux plus élevés d’amplitude et de fréquence des ondes de contraction de l’utérus par rapport aux autres groupes EA [[31]].
Commentaires sur le cas clinique
En fonction des données issues des ECR et de l’électroacupuncture expérimentale, il s’avère que l’EA est plus efficace dans la maturation du col puis l’induction des contractions utérines que l’acupuncture seule. La fréquence de stimulation doit être basse, entre 2 et 50Hz, soit en fréquence unique de 2Hz ou alternée à celle de 50Hz. Le choix des points est également important. Toutes les études ont un minimum de points communs qui sont taichong (3F), hegu (4GI), sanyinjiao (6Rt), ciliao 32V. On peut ajouter 3VC (zhongji) et 4VC (guanyuan) qui favoriseraient davantage la maturation du col. Ainsi selon la médecine chinoise, 3VC qui est le point mu de Vessie et également point de croisement avec les Méridiens de Rein, Foie et Rate-Pancréas (comme le 4VC), draine l’Humidité et fortifie la déficience du yang qi, de l’énergie originelle yuan qi et du Rein tout comme le 4VC. Il est à utiliser effectivement avec le 4VC, lui même point mu d’Intestin Grêle et point majeur dans les dystocies du col, favorisant ainsi la maturation du col.
Conclusion
A partir des données issues de l’acupuncture factuelle, L’EA ou la neurostimulation électrique transcutanée appliquée aux points d’acupuncture est plus efficace que l’acupuncture manuelle dans les nausées, vomissements, dans la maturation du col utérin et l’induction du travail chez la femme enceinte. Aucune complication obstétricale n’a été décelée, confirmant l’innocuité et la sécurité de l’EA. Et comme pour les algies [1], l’utilisation de l’acupuncture chez la femme enceinte peut être proposée avec un grade B de présomption scientifique de niveau 2 de preuves selon les recommandations de la Haute Autorité de Santé française (HAS) [[32]] en attendant d’avoir un ECR de très grande qualité méthodologique.
Notes
[a]. La distension excessive de l’estomac active des récepteurs à l’étirement qui déclenchent un réflexe vaso-vagal provoquant la relaxation du sphincter inférieur de l’œsophage.
[b]. L’oxyde nitrique (NO) est le neuro-transmetteur non adrénergique et non cholinergique. Il permet la relaxation des muscles lisses de l’appareil gastro-intestinal.
[c]. Bishop EH. Pelvic scoring for elective induction. Obstet Gynecol. 1964;24:266-8.
[d]. HAS. Déclenchement artificiel du travail à partir de 37 semaines d’aménorrhée. 2008. [cité le 16/08/2016]. Available from URL : http://www.has-sante.fr/portail/upload/docs/application/pdf/declenchement_artificiel_du_travail_-_argumentaire.pdf
[d]. Acupuncture sham par aiguilles placebo de type Park
[f]. Protocole d’acupuncture utilisant 4GI, 36E, 3R (taixi), 60V (kunlun), 31V, 32V, 20VG (baihui), 6Rt.
Références
[1]. Stéphan JM. Electroacupuncture chez la femme enceinte – abord pratique et théorique dans les algies. Acupuncture & Moxibustion. 2016;15(2):130-138.
[2]. Rosen T, de Veciana M, Miller HS, Stewart L, Rebarber A, Slotnick RN. A randomized controlled trial of nerve stimulation for relief of nausea and vomiting in pregnancy. Obstet Gynecol. 2003;102(1):129-35.
[3]. Matthews A, Haas DM, O’Mathúna DP, Dowswell T. Interventions for nausea and vomiting in early pregnancy. Cochrane Database Syst Rev. 2015;9:CD007575.
[4]. Ezzo JM, Richardson MA, Vickers A, Allen C, Dibble SL, Issell BF, Lao L, Pearl M, Ramirez G, Roscoe J, Shen J, Shivnan JC, Streitberger K, Treish I, Zhang G. Acupuncture-point stimulation for chemotherapy-induced nausea or vomiting. Cochrane Database Syst Rev. 2006;(2):CD002285.
[5]. Shen J, Wenger N, Glaspy J, Hays RD, Albert PS, Choi C, Shekelle PG. Electroacupuncture for control of myeloablative chemotherapy-induced emesis: A randomized controlled trial. JAMA. 2000;284(21):2755-61.
[6]. Yu Z, Cao X, Xia Y, Ren B, Feng H, Wang Y, Jiang J, Xu B. Electroacupuncture Stimulation at CV12 Inhibits Gastric Motility via TRPV1 Receptor. Evid Based Complement Alternat Med. 2013;2013:294789.
[7]. Tatewaki M, Strickland C, Fukuda H, Tsuchida D, Hoshino E, Pappas TN,Takahashi T. Effects of acupuncture on vasopressin-induced emesis in conscious dogs. Am J Physiol Regul Integr Comp Physiol. 2005;288(2):R401-8.
[8]. Shiotani A, Tatewaki M, Hoshino E, Takahashi T. Effects of electroacupuncture on gastric myoelectrical activity in healthy humans. Neurogastroenterol Motil. 2004;16(3):293-8.
[9]. Zou D, Chen WH, Iwakiri K, Rigda R, Tippett M, Holloway RH. Inhibition of transient lower esophageal sphincter relaxations by electrical acupoint stimulation. Am J Physiol Gastrointest Liver Physiol. 2005;289(2):G197-201.
[10]. Wang C, Zhou DF, Shuai XW, Liu JX, Xie PY. Effects and mechanisms of electroacupuncture at PC6 on frequency of transient lower esophageal sphincter relaxation in cats. World J Gastroenterol. 2007;13(36):4873-80.
[11]. Yang J, Wang C. [Electroacupuncture at « Zusanli »(ST 36) can inhibit frequencies of transient lower esophageal sphincter relaxation induced by gastric distention in cats]. Zhen Ci Yan Jiu. 2011;36(6):423-7.
[12]. Wang C, Chen X, Xie PY. Electroacupuncture at PC6 or ST36 Influences the Effect of Tacrine on the Motility of Esophagus. Evid Based Complement Alternat Med. 2014;2014:263489.
[13]. Smith CA, Crowther CA. Acupuncture for induction of labour. Cochrane Database of Systematic Reviews 2004, Issue 1.
[14]. Gaudernack LC, Forbord S, Hole E. Acupuncture administered after spontaneous rupture of membranes at term significantly reduces the length of birth and use of oxytocin. A randomized controlled trial. Acta Obstet Gynecol Scand. 2006;85(11):1348-53.
[15]. Harper TC, Coeytaux RR, Chen W, Campbell K, Kaufman JS, Moise KJ, et al. A randomized controlled trial of acupuncture for initiation of labor in nulliparous women. J Matern Fetal Neonatal Med. 2006 Aug;19(8):465-70.
[16]. Rabl M, Ahner R, Bitschnau M, Zeisler H, Husslein P. Acupuncture for cervical ripening and induction of labor at term–a randomized controlled trial. Wien Klin Wochenschr. 2001;113(23-24):942-6.
[17]. Stéphan JM. L’acupuncture autour de la naissance : bases scientifiques et état des lieux dans les versions, induction du travail et maturation du col. Acupuncture & Moxibustion. 2009;8(3):157-166.
[18]. Smith CA, Crowther CA, Grant SJ. Acupuncture for induction of labour. Cochrane Database Syst Rev. 2013;(8):CD002962.
[19]. Modlock J, Nielsen BB, Uldbjerg N. Acupuncture for the induction of labour: a double-blind randomised controlled study. BJOG. 2010;117(10):1255-61.
[20]. Gribel GP, Coca-Velarde LG, Moreira de Sá RA. Electroacupuncture for cervical ripening prior to labor induction: a randomized clinical trial. Arch Gynecol Obstet. 2011;283(6):1233-8.
[21]. Tremeau ML, Fontanie-Ravier P, Teurnier F, Demouzon J. [Protocol of cervical maturation by acupuncture]. J Gynecol Obstet Biol Reprod (Paris). 1992;21(4):375-80.
[22]. Asher GN, Coeytaux RR, Chen W, Reilly AC, Loh YL, Harper TC. Acupuncture to initiate labor (Acumoms 2): a randomized, sham-controlled clinical trial. J Matern Fetal Neonatal Med. 2009;22(10):843-8.
[23]. Gaudet LM, Dyzak R, Aung SK, Smith GN. Effectiveness of acupuncture for the initiation of labour at term: a pilot randomized controlled trial. J Obstet Gynaecol Can. 2008;30(12):1118-23.
[24]. Mackenzie IZ, Xu J, Cusick C, Midwinter-Morten H, Meacher H, Mollison J, Brock M. Acupuncture for pain relief during induced labour in nulliparae: a randomised controlled study. BJOG. 2011;118(4):440-7.
[25]. Selmer-Olsen T, Lydersen S, Morkved S. Does acupuncture used in nulliparous women reduce time from prelabour rupture of membranes at term to active phase of labour? A randomised controlled trial. Acta Obstet Gynecol Scand. 2007;86(12):1447-52.
[26]. Smith CA, Crowther CA, Collins CT, Coyle ME. Acupuncture to induce labor: a randomized controlled trial. Obstet Gynecol. 2008;112(5):1067-74.
[27]. Ajori L, Nazari L, Eliaspour D. Effects of acupuncture for initiation of labor: a double-blind randomized sham-controlled trial. Arch Gynecol Obstet. 2013;287(5):887-91.
[28]. Andersen BB, Knudsen B, Lyndrup J, Fælling AE, Illum D, Johansen M, Borgen A, Jager H, Bjerre C, Secher NJ. Acupuncture and/or sweeping of the fetal membranes before induction of labor: a prospective, randomized, controlled trial. J Perinat Med. 2013;41(5):555-60.
[29]. Neri I, Monari F, Midwife CS, Facchinetti F. Acupuncture in post-date pregnancy: a pilot study. J Matern Fetal Neonatal Med. 2014 ;27(9):874-8.
[30]. Dunn PA, Rogers D, Halford K. Transcutaneous electrical nerve stimulation at acupuncture points in the induction of uterine contractions. Obstet Gynecol. 1989;73(2):286-90.
[31]. Chen YR, Yuan HW, Shu FZ, Ren XX, Ma LX, Liu RQ, Zhu J. [Effects of electroacupuncture at points of a traditional acupuncture formula that induces labor with different parameters on uterine contractility in late-stage pregnant rats]. Zhong Xi Yi Jie He Xue Bao. 2011 Jan;9(1):91-9.
[32]. HAS. Niveau de preuve et gradation des recommandations de bonne pratique – État des lieux. 2013. [cité le 11/09/2016]. Available from URL : http://www.has-sante.fr/portail/upload/docs/application/pdf/2013-06/etat_des_lieux_niveau_preuve_gradation.pdf.



