Résumé : Dès la fin des années 1970, les mécanismes neurophysiologiques de l’électroacupuncture dans les algies commencent à être dévoilés. Les peptides opioïdes endogènes (endorphines, enképhalines, dynorphines, endomorphines) et leurs récepteurs µ, κ , δ, le système anti-opioïde (nociceptine, CCK-8…), le glutamate et ses récepteurs ionotropiques AMPA, KA, NMDA et enfin le système inhibiteur descendant faisant intervenir sérotonine et cathécolamines sont les différents mécanismes que l’électroacupuncture active en fonction de la fréquence électrique. Par l’exploration expérimentale chez l’animal, mais aussi chez l’homme par les techniques d’imagerie (IRM fonctionnelle, tomographie par émission de positons), on sait maintenant que les aires cérébrales activées correspondent à celles impliquées dans la douleur (par activation ou inactivation) : systèmes hypothalamique, limbique et paralimbique, mais aussi tronc cérébral et corne dorsale de la moelle épinière. Mots-clés : électroacupuncture – endorphine – dynorphine – anti-opioïdes – CCK-8 – système inhibiteur descendant – IRMf – TEP – système limbique.
Summary: In the late 1970’s, the neurophysiological mechanisms of electroacupuncture pain in the beginning to be unveiled. The endogenous opioid peptides (endorphins, enkephalins, dynorphins, endomorphins) and theirs µ, κ , δ receptors, antiopioids system (nociceptin, CCK-8…), glutamate and his ionotropics AMPA, KA, NMDA receptors then finally the descending inhibitory system involving serotonin and cathecolamins are the different mechanisms that electroacupuncture active as a function of frequency electric. By exploring experimental animals, but in humans through imaging techniques (functional MRI, positron emission tomography), we now know that the brain areas activated correspond to those involved in pain (activation or inactivation): hypothalamic and limbic systems, but also brain stem and dorsal horn of the spinal cord. Keywords: electroacupuncture – endorphin – dynorphin – antiopioid – descending inhibitory system – CCK-8 – fMRI – PET.
Les molécules informationnelles, substances chimiques produites par une cellule vivante pour transmettre un signal à une autre cellule qui le reçoit à son tour par un récepteur spécifique, sont à la base de l’Acupuncture. De nombreuses molécules sont issues des voies de la mécanotransduction ou de la transduction [[1],[2]] et certaines d’entre-elles interviennent dans l’obtention des effets de l’acupuncture et/ou de l’électroacupuncture (EA). La démonstration de récepteurs spécifiques aux alcaloïdes au niveau cérébral a permis de découvrir des ligands naturels endogènes, les opioïdes actifs à structure peptidique (neuropeptides) qui jouent le rôle de médiateurs ou de neuromodulateurs. Le glutamate, un autre neurotransmetteur excitant majeur et ses récepteurs ionotropiques NMDA, AMPA et kainate (KA) interviennent aussi dans la modulation des algies par électroacupuncture ainsi que le système inhibiteur descendant sérotoninergique et noradrénergique. Grâce à l’imagerie cérébrale, il est possible d’observer les différentes cibles du système nerveux central intervenant dans l’électroacupuncture.
Rappels neurophysiologiques de la douleur
Les stimulations nociceptives activent les terminaisons libres des nocicepteurs C ou Aδ. La physiologie de la douleur aiguë puis chronique a beaucoup profité des progrès de la biologie moléculaire qui a permis de découvrir une « soupe » inflammatoire complexe périphérique : ATP et récepteur purinergique P2X3, récepteur vanilloïde sensible à la chaleur, bradykinine, substance P et CGRP (calcitonin gene-related peptide), nerve growth factor etc.. Les acides aminés excitateurs comme le glutamate, sérotonine, peptides opioïdes etc.. vont moduler la transmission de cette douleur de la première synapse à l’arrivée des fibres périphériques dans la moelle. Puis un relais s’effectuera vers des cibles supraspinales : bulbe, mésencéphale, thalamus, système limbique et cortex. Le caractère douloureux dépendra d’une balance entre des influx excitateurs et inhibiteurs comme le système inhibiteur descendant issu du tronc cérébral. Nous allons découvrir les principales molécules agissantes de l’électroacupuncture.
Les peptides opioïdes endogènes
Localisation de la synthèse des neuropeptides
Ces endorphines sont synthétisées à deux niveaux :
– dans le système nerveux central (SNC) : corne dorsale de la moelle épinière, substance réticulée (noyaux du raphé médian et para-médian et les noyaux giganto-cellulaires), les ganglions de la base (noyau caudé, putamen, noyau sous-thalamique, pars lateralis, pallidum), mésencéphale (substance grise périaqueducale, noyau inter-pédonculaire, noyau parabrachial, noyau du faisceau solitaire, substance noire), amygdale, hippocampe, diencéphale (hypothalamus, lobe antérieur de l’hypophyse, thalamus), cervelet ; cortex cérébral (sauf occipital) ;
– dans le système nerveux périphérique : neurones du ganglion mésentérique, neurones intrinsèques ou cellules endocrines de l’intestin, pancréas, cœur, poumons, médullo-surrénales et les organes de la reproduction.
Les différents opioïdes et leurs récepteurs
Trois familles de neuropeptides se distinguent par leur précurseur protéique spécifique dont elles sont issues par protéolyse : la proopiomélanocortine (POMC), la préproenképhaline (PPE) et la préprodynorphine (PPD). Les endorphines résultantes de la protéolyse de ces précurseurs préotéiques ont la même séquence amino-terminale identique Tyr-Gly-Gly-Phe-Met (Met-enképhaline) ou Tyr-Gly-Gly-Phe-Leu (Leu-enképhaline) (figure 1). La protéolyse de la POMC produit entre autres neuropeptides, l’hormone adénocorticotrope (ACTH), l’alpha MSH, la β-lipotrophine, les bêta endorphines. La PPE engendre les Met-enképhalines et peptides voisins ; la PPD les dynorphines et les néoendorphines. On a isolé plus de 20 peptides issus de ces trois familles avec une exception, les endomorphines 1 et 2 qui ne possèdent pas la séquence pentapeptique, mais quatre acides aminés [[3]]. Leur rôle comme neurotransmetteur ou neuromodulateur est très probable mais incomplètement élucidé.
Figure 1. Structure des enképhalines.
Plusieurs récepteurs opioïdes ont, eux aussi, été identifiés et différenciés.
Dans le SNC, trois classes principales sont distinguées : µ (mu/OP3/MOR) avec les sous types μ1, μ2, κ (kappa/OP2/KOR) avec les sous-types κ1, κ2, κ3, δ (delta/OP1/DOR). Certaines observations suggèrent l’existence d’autres types de récepteurs : epsilon (ε), zêta (ξ) et lambda (λ). Une substance opioïde donnée peut interagir avec les trois récepteurs différents et se comporter, pour l’un, comme un agoniste, pour l’autre, comme un agoniste partiel ou même comme un antagoniste. Pour cette raison, il peut exister des différences d’effets entre les opioïdes. Les morphiniques agissent par l’activation d’une protéine G couplée aux récepteurs µ, κ , δ par le mécanisme de transduction [[4]].
Les enképhalines par exemple, activent divers types de récepteurs du SNC, parmi lesquels les récepteurs μ (mu), μ1, μ2, et κ1, κ2, κ3, sont les mieux décrits. L’activation des récepteurs µ, appelés aussi OP3 (Opioid receptor) engendre les effets morphiniques des endorphines et entraîne analgésie, dépression respiratoire, constipation, dépendance, myosis, hypothermie. Le récepteur δ est plus spécifique des enképhalines qui sont co-sécrétées avec les catécholamines par la médullosurrénale entraînant l’euphorie et l’analgésie également. L’activation des récepteurs κ qui présentent une affinité particulière pour les dynorphines, provoque analgésie, sédation, myosis. L’analgésie par les morphiniques peut provenir de l’activation des récepteurs μ et κ et la dépendance par l’activation des récepteurs μ (voir tableau résumé I).
Tableau I : Classification des opioïdes et effets de leurs différents récepteurs.
Le SNC synthétise et libère des molécules, que l’on peut qualifier d’anti-opioïdes qui sont libérées à la suite de la prise de toute substance capable de stimuler les récepteurs des opioïdes. Il s’agit des réseaux neuronaux capables de s’opposer aux actions des opioïdes et qui peuvent expliquer en partie la tolérance et la dépendance. Récemment un nouveau type de récepteur a été ainsi identifié et appelé récepteur-orphelin aux opiacés, ORL1 (opioid receptor-like / OP4) dont l’agoniste est la nociceptine. Les principales molécules anti-opioïdes sont la cholécystokinine-8 (CCK-8), la neuropeptide FF (NPFF), la melanocyte inhibiting factor (MIF) et la nociceptine appelée auparavant orphanine FQ, protéine neuropeptide de 17 acides aminés ayant des similarités avec la dynorphine A (voir tableau II).
Tableau II : Classification des anti-opioïdes et effets de leurs différents récepteurs.
Classe
Récepteurs
Effets pharmacologiques
Localisation de la synthèse
nociceptine ou orphanine FQ
ORL1
Pronociceptif et antiopioïde au niveau supraspinalAnalgésique / antinociceptive au niveau spinalanxiolytique
très faible affinité pour les récepteurs μ, δ et k
Pronociceptif
Système nerveux central
melanocyte inhibiting factor (MIF)
Agonistes partiels des récepteurs de type μ
Pronociceptif
Système nerveux central
Système inhibiteur descendant supraspinal
Il existe deux types de contrôle inhibiteurs descendants.
Contrôle descendant issu du tronc cérébral et déclenché par des stimulations cérébrales
La stimulation chez l’animal du raphé, du bulbe (région bulbaire rostro-ventrale comprenant le noyau raphé magnus, le noyau giganto-cellulaire, le noyau réticulé latéral du tractus solitaire), du pont et du mésencéphale (substance grise périaqueducale) entraîne une analgésie par blocage des entrées nociceptives dans le système nerveux central. Les neurones du raphé sont sérotoninergiques et se projettent dans la partie dorsale du faisceau latéral de la moelle (funiculus dorsal), mais sont aussi interconnectés au locus coeruleus, structure noradrénergique. Il y a donc aussi une implication d’un contingent de fibres noradrénergiques mais aussi des substances opioïdes
Contrôle inhibiteur descendant déclenché par des stimulations nociceptives (CIDN)
Ce contrôle est déclenché par stimulation nociceptive (CIDN) périphérique. La structure impliquée est la réticulée bulbaire. Les neuromédiateurs sont endorphiniques et sérotoninergiques.
Le glutamate et ses principaux récepteurs
Le glutamate
Il intervient comme neurotransmetteur excitateur qui libéré dans l’espace synaptique, à partir des terminaisons neuronales, se fixe sur ses récepteurs postsynaptiques dont l’activation induit la dépolarisation du neurone cible. Cependant, si la régulation de cette transmission synaptique est altérée et l’activation des récepteurs prolongée, il peut y avoir maintien d’une dépolarisation accrue conduisant à la mort cellulaire. On a montré que l’injection par voie systémique de glutamate ou d’autres acides aminés excitateurs (AAE) à des animaux immatures entraînait des dégénérescences dans des aires du cerveau qui ne sont pas protégées par la barrière hémato-encéphalique. Depuis, de nombreuses études ont mis en évidence la toxicité des AAE, ou excitotoxicité. Le glutamate en trop grande concentration a été ainsi mis en cause dans l’étiologie de nombreuses pathologies du système nerveux central : lésions traumatiques du SNC, maladies neurodégénératives aiguës, maladies neurodégénératives inflammatoires (scléroses multiples), maladies neurodégénératives chroniques (Alzheimer, Chorée de Huntington, SLA).
Les récepteurs ionotropiques du glutamate
Le glutamate agit sur trois récepteurs canaux distincts dénommés par le nom de leur agoniste le plus sélectif : les récepteurs N-méthyl-D-aspartate (NMDA), kainate (KA) et a-amino-3-hydroxy-5-métyl-4-isoxazolepropionate (AMPA). Ces trois récepteurs sont des récepteurs ionotropiques capables de transmettre instantanément un message au neurone cible par modification du potentiel de la membrane post-synaptique en quelques millièmes de secondes.
On a mis en évidence deux familles de récepteurs, respectivement AMPA et kainate (KA) à la fin des années 1970. Les sous-unités AMPA et kainate peuvent être colocalisées au sein d’un même neurone mais ne peuvent s’assembler entre-elles. Les différentes sous-unités des récepteurs AMPA/KA sont abondamment exprimées dans l’ensemble du SNC. Les récepteurs NMDA sont quant à eux mis en jeu dans de nombreux mécanismes physiologiques comme la différenciation neuronale et la formation des connections synaptiques au cours du développement. Chez l’adulte, le récepteur NMDA est impliqué dans l’apprentissage et la mémoire à court terme. Les récepteurs au NMDA jouent aussi un rôle important dans différents processus physiologiques en augmentant la transmission du processus douloureux. La kétamine est un antagoniste des récepteurs au NMDA et son injection produit une analgésie puissante.
Neurophysiologie de l’action de l’électroacupuncture
Electroacupuncture analgésique expérimentale
Découverte des opioïdes
Le premier travail expérimental sur l’analgésie acupuncturale fut réalisé dans les années 1970 chez 60 étudiants en médecine volontaires [[5]]. La douleur fut induite par ionophorèse potassique à travers la peau. Le seuil à la douleur fut mesuré après insertion et manipulation pendant 50 minutes des aiguilles d’acupuncture au GI4 et ES36. Dans le groupe contrôle, 10 mg de morphine en intra-musculaire augmente le seuil de douleur en moyenne de 80 à 90%. Dans le groupe acupuncture, on observa de façon identique une augmentation graduelle du seuil à la douleur avec un pic au bout de 20 à 40 minutes après l’insertion de l’aiguille. En cas d’injection de procaïne, anesthésique local, l’effet de l’acupuncture est annulé, ce qui suggère que l’effet nécessitait des récepteurs sensoriels intacts.
En 1974, la même équipe présupposant le rôle humoral de neurotransmetteurs, montre que l’analgésie induite par acupuncture peut être transmise d’un lapin à un autre par transfusion du liquide céphalo-rachidien (LCR) [[6]]. Tous ces travaux sont conduits un peu avant la découverte des endorphines.
Lorsque les premières endorphines sont découvertes en 1975, nombreux sont ceux qui font l’hypothèse que ces substances sont les médiateurs responsables des effets de l’analgésie acupuncturale.
En 1976, Pomeranz et coll. [[7]] montrent chez la souris que la naloxone bloque l’effet analgésique de l’acupuncture, ce qui implique le rôle des endorphines. Chez l’homme, Sjolund et coll. [[8]] en 1977 démontrent que l’induction de l’analgésie par EA pendant 30 mn s’accompagne d’une augmentation dans le LCR du niveau de bêta-endorphines issues de la région hypothalamique et confirment que l’administration d’un antagoniste opiacé, la naloxone inhibe totalement l’analgésie. Mayer et coll. rapportent aussi que l’analgésie par acupuncture est réversible 5 mn après l’injection de la naloxone [[9]]. Clement-Jones et coll. en 1980 objectivent chez 10 volontaires présentant des douleurs chroniques une élévation dans le LCR du taux des bêta-endorphines après 30 mn d’EA à basse fréquence (2-3 Hz) alors que le niveau de la met-enképhaline n’est pas augmenté. Cependant, ils supposaient déjà que différents mécanismes étaient impliqués, fonction de la fréquence de stimulation [[10]].
En 1992, Chen et Han montrent que l’analgésie produite par l’EA est régulée par trois types de récepteurs opioïdes [[11],[12]]. Ainsi, l’EA à 2 Hz active les récepteurs μ et δ ; celle à 100 Hz, les récepteurs κ. Mais mieux, l’EA à 15 Hz produit une activation des trois sortes de récepteurs chez le rat [[13]].
Chez le rat anesthésié, la stimulation électrique à basse fréquence (4 Hz) de zusanli (ES36) entraîne une expression de c-fos dans le lobe antérieur de la glande hypophysaire, aussi bien qu’au niveau des noyaux hypothalamiques arqués et autres voisins [[14]]. Les mêmes auteurs ont montré dans une autre étude chez les rats anesthésiés au pentobarbital et subissant une stimulation nociceptive thermale que l’EA (4 Hz sur zusanli) entraînait une augmentation de la localisation de l’expression de c-fos dans le noyau hypothalamique médio-basal et arqué, et dans le noyau paraventriculaire hypothalamique. On retrouvait aussi une élévation de l’hormone adrénocorticotrope (ACTH) et des bêta endorphines plasmatiques. Cela suggère que l’axe hypothalamo hypophyso surrénalien est donc activé lors de l’action de l’électroacupuncture [[15],[16]]. Néanmoins, l’étude de Yang et coll. objective l’implication directe des neurones du noyau supraoptique hypothalamique (SON) dans l’analgésie par EA (36ES 10/20 Hz alternés, 30 mn). L’ocytocine et la vasopressine, sécrétées par le SON, sont connues pour augmenter le seuil à la douleur. Cependant, les auteurs ont montré qu’il fallait distinguer les effets liés au passage des fibres nerveuses reliées aux régions extra-hypothalamiques (substance périaqueducale, noyau raphé magnus, amygdale, locus coerulus etc..), des effets liés aux neurones propres du SON (neurones magnocellulaires sécréteurs de vasopressine et d’ocytocine) et aux dendrites libérant les neuropeptides (modulés aussi par l’ocytocine) [[17]].
Par des travaux d’acupuncture expérimentale sur les animaux, rats, lapins, des auteurs à leur tour démontrent la libération des endorphines par électroacupuncture au niveau du système limbique : amygdale, hippocampe, thalamus, noyau accumbens etc.. ; du bulbe : substance réticulée (les noyaux raphé magnus, giganto-cellulaire, noyau ambigu) ; hypothalamus ; mésencéphale (substance grise périaqueducale) ; corne postérieure de la moelle épinière (funiculus postéro-latéral) etc.. et leur inhibition par la naloxone ou la naltrexone (inhibiteurs opioïdes) [18-28].
Contrôles inhibiteurs descendants
Autre neuromédiateur, la sérotonine ou 5-hydroxytryptamine joue un rôle important lors d’une stimulation par EA et interviendrait, entre autres, dans l’un des mécanismes de l’analgésie par stimulation du noyau raphé dorsal [29-32].
Takagi et coll. ont déterminé chez le lapin que les récepteurs, localisés au niveau présynaptique et postsynaptique, et libérés dans l’EA (36 ES, 2Hz, 40mn) étaient le 5-HT1 (excepté le 5-HT1A) ; le 5-HT2 (excepté le 5-HT2A) et le récepteur 5-HT3 [[33]]. L’EA à 2Hz stimule aussi l’expression du récepteur 5-HT3a [[34]].
En 2007, Li et coll. montrent que l’EA (10 Hz à 3mA) appliquée pendant 20mn dans un modèle d’algie par injection d’adjuvant de Freund sur la patte de rat inhibe la transmission de la nociception et de l’hyperalgie en activant les neurones supraspinaux qui se projettent sur la moelle épinière par le système inhibiteur descendant. L’EA active la sérotonine et les catécholamines des neurones du noyau raphé magnus et du locus coeruleus dont l’expression c-fos est significativement augmentée [[35]].
Récemment, il a été démontré que l’analgésie par EA était aussi médiée par les récepteurs α2 adrénergiques de la moelle épinière. Ceci a été observé de manière statistiquement significative sur un modèle d’entorse de cheville chez le rat traité par EA à 100 Hz (largeur d’impulsion carrée de 1 ms) sur TR6 (zhigou) pendant 30 mn versus 4GI (hegou) [[36]]. Plus précisément, Kim et coll. en 2008 observent que la suppression des effets de l’EA à basse fréquence (1 Hz) sur un modèle de douleur inflammatoire carragénine induite sur la patte de rat est médiée par les ganglions sympathiques post-synaptiques. La haute fréquence (120 Hz) intervient quant à elle, par le système médullaire sympatho-surrénalien. Ceci est démontré par le fait que la surrénalectomie diminue l’action de l’EA à 120 Hz sans affecter la basse fréquence et que le traitement pré-thérapeutique avec un antagoniste des récepteurs à la corticostérone n’intervient pas dans l’efficacité de l’EA à 2 ou 120 Hz. D’autre part, l’administration de la 6-hydroxydopamine (neurotoxine des terminaisons nerveuses sympathiques périphériques) bloque sélectivement l’EA (2 Hz). Le propranolol (antagoniste des récepteurs bêta-adrénergiques) abolit complètement à la fois l’EA haute et basse fréquence [[37]].
Récepteur ionotropique NMDA
L’électroacupuncture (36ES, 6RP) à la fréquence de 2 Hz agit aussi dans les algies en diminuant la réponse à l’inflammation locale par l’intermédiaire de la modulation de l’expression des récepteurs ionotropes au glutamate et en particulier le récepteur au N-méthyl-D-aspartate (NMDA) dans la corne dorsale de la moelle épinière [[38],[39]]. Wang et coll. vont confirmer en 2006 le rôle de la modulation de l’expression des récepteurs de la NMDA par EA (4 Hz/16 Hz alternativement pendant 30 mn : 30VB huantiao et 34VB yanglingquan) sur un modèle de rat hyperalgique [[40]].
Un stimulus nociceptif ou une lésion aiguë d’un nerf engendre une potentialisation à long terme des potentiels évoqués des fibres C de la corne dorsale de la moelle. L’EA à basse fréquence (2 Hz de 1 à 3mA par incrément de 1 mA toutes les 10 mn) appliquée sur ES36 et RA6 induit une dépression à long terme de ces mêmes potentiels évoqués chez le rat ayant une ligature du nerf rachidien de L5-L6. Cet effet est bloqué par l’antagoniste des récepteurs de la N-méthyl-d-acide aspartique (NMDA), le MK-801 et par l’antagoniste des récepteurs opioïdes, la naloxone. En revanche, l’EA à haute fréquence (100 Hz), qui n’est pas efficace dans le traitement des douleurs neuropathiques (encore appelée douleur par désafférentation), induit une potentialisation à long terme des potentiels évoqués des fibres C de la corne dorsale de la moelle chez le rat avec ligature du nerf rachidien. Contrairement à l’EA à 2 Hz, l’EA à 100 Hz est dépendante des systèmes inhibiteurs sérotoninergiques et gabanergiques. L’EA à 2 Hz intervient dans la plasticité synaptique rachidienne par une dépression à long terme de la transmission synaptique au niveau de la corne dorsale de la moelle épinière. Les effets à long terme de l’antinociception s’expliqueraient par l’induction des récepteurs NMDA à dépression à long terme via l’activation du système des peptides opioïdes endogènes [[41]].
En 2008, à nouveau Ryu et coll. objectivent que l’EA (2Hz, 1,0 mA au 36ES et 6RP 30 mn) entraîne chez le rat une modulation de la phosphorylation des récepteurs NMDA (NR1 et NR2B) au niveau de la moelle épinière [[42]]. Le tableau III récapitule les principaux travaux d’électroacupuncture expérimentale et d’imagerie avec les résultats observés.
Tableau III. Récapitulatif des principaux travaux d’électroacupuncture expérimentale et d’imagerie.
Auteur (année)
Caractéristiques de l’électroacupuncture
Localisation cérébrale : visualisation par expression c-fos / ac anti-récepteurs stimulés / imagerie fonctionnelle
100 Hz (8 à 15 mA)36ES (zusanli) et 6RP (sanyinjiao)chez 48 volontaires sains
Activation : lobule pariétal inférieur controlatéral, le cortex cingulaire antérieur ipsilatéral et le noyau accumbens,
Désactivation :amygdale controlatérale
Choi (2005)[39]
2 Hz, 15 Hz et 120 Hz : 36ES, 6RPchez le rat pendant 30j par intervalle de 3 jours
Corne dorsale de la moelle épinière : modulation des récepteurs NMDA (type NR1, NR-2A, GlucR-1, GluR-2/3) pour tous les fréquences
Diminution des récepteurs ionotropiques NMDA (NR1 et NR-2A)
Napadow (2005)[46]
36ES (zusanli)courant continu de 0,7 à 3,6mA ;7 mn13 volontaires sains
2 Hz
Visualisation par IRM fonctionnelle
Intervention du système limbique
Activation :insula antérieurcortex cingulaire antérieur médial Désactivationamygdale, hippocampe antérieur, cortex cingulaire rétrosplénial (BA29 et BA30), le cortex cingulaire subgenual, le cortex ventromédial préfrontal, les lobes frontaux et temporaux.
Aire du raphé du pont.
100 Hz
Li (2007)[35]
10 Hz (3 mA) 20mn30VBAlgie induite par adjuvant de Freund chez rat
Inhibition expression c-fos-couches lame I et II de la corne dorsale de la moelle épinièrestimulation-noyau raphé magnus- locus ceruleus
intervention du système inhibiteur descendant : sérotonine et les catécholamines des neurones du noyau raphé magnus et du locus coeruleus
Imagerie électroacupuncturale analgésique
L’étude de Wu et coll. de 2002 [[43]] a étudié la réaction cérébrale par IRMf suite à la stimulation électrique du point VB34 (yanglinquan) utilisé en analgésie chez 15 volontaires sains. Ils objectivaient que l’EA vraie à 4 Hz activait de manière statistiquement significative par rapport au groupe placebo (EA « sham » appliquée sur des non-points d’acupuncture) l’hypothalamus, l’aire primaire somatosensorielle ou somatosentivive (S1), le cortex moteur et désactivait le segment rostral du cortex cingulaire antérieur qui est impliqué dans la « réponse émotive » à la douleur. Ils concluaient que les systèmes limbique et hypothalamique étaient modulés par l’électroacupuncture.
Zhang et coll. en 2003 vont démontrer également que l’effet analgésique est modulé à la fois par l’aspect sensoriel et émotionnel du processus douloureux en réponse à la stimulation électrique des points d’acupuncture. Chez huit volontaires sains chez qui on applique un stimulus de chaleur (18°C) et de froid (2°C), l’électroacupuncture engendre une activation de l’aire bilatérale somatosensorielle secondaire (S2), du cortex préfrontal médial, de l’aire de Brodman 32 (BA32) et une désactivation de l’aire S1 controlatérale, de la BA7 et BA24 (gyrus cingulaire antérieur) [[44]].
Les mêmes auteurs [[45]] ont étudié par IRM fonctionnelle les mécanismes possibles à la base de la spécificité de fréquence. Dans les deux cas, il y a des niveaux d’activation positives dans les zones S2 bilatérales, dans l’insula, dans le cortex cingulaire antérieur controlatéral et le thalamus en rapport avec l’effet analgésique. A la fréquence de 2 Hz, des corrélations positives ont été observées dans l’aire motrice primaire controlatérale (gyrus précentral), l’aire motrice supplémentaire et le gyrus temporal supérieur ipsilatéral, tandis que des corrélations négatives ont été retrouvées dans l’hippocampe bilatéral. A la fréquence de 100 Hz, des activations positives ont été observées dans le lobule pariétal inférieur controlatéral, le cortex cingulaire antérieur ipsilatéral et le noyau accumbens, tandis qu’une corrélation négative a été détectée dans l’amygdale controlatérale. Ces résultats montrent que l’analgésie en rapport avec l’électroacupuncture à fréquences basses et hautes nécessite la médiation de différents réseaux cérébraux plus ou moins entremêlés.
Napadow et coll. objectivent que l’EA à 2Hz et à 100 Hz ainsi que l’acupuncture manuelle produisent une activation du signal BOLD de l’IRM fonctionnelle au niveau de l’insula antérieure et une désactivation des structures limbiques et paralimbiques. Augmentation aussi du signal BOLD dans le cortex cingulaire antérieur médial à la fois pour l’EA haute et basse fréquence, toutefois seule l’EA à basse fréquence (2Hz) produit une activation de l’aire du raphé du pont. Tous ces résultats confirment l’hypothèse que le système limbique est un élément central de l’analgésie électroacupuncturale [[46]].
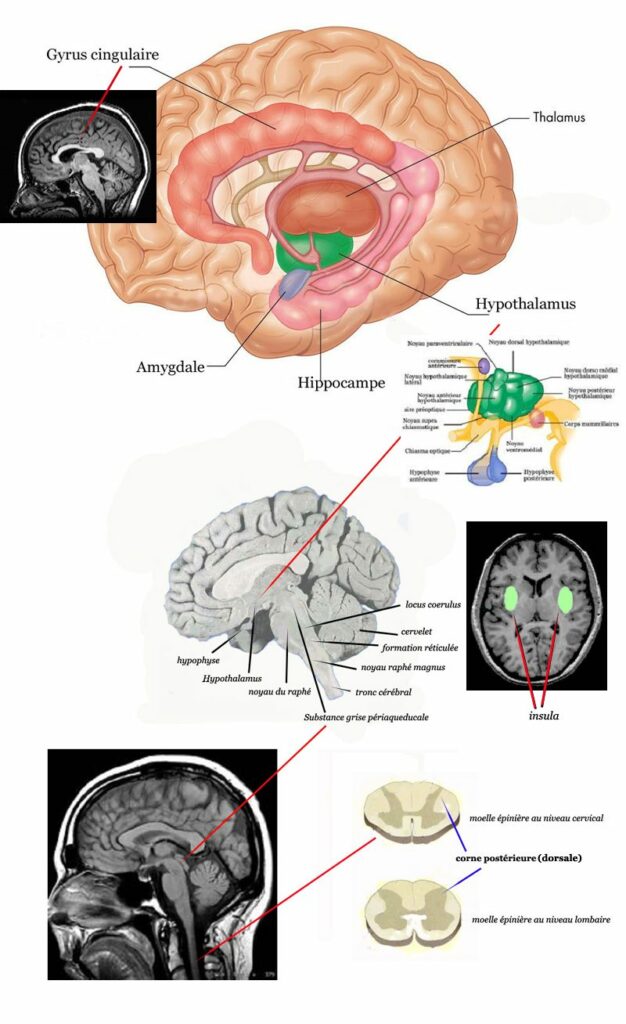
L’étude de l’équipe japonaise de Maenaka s’intéresse à la suppression de la douleur chaleur-induite chez trois singes rhésus et à sa visualisation cérébrale par utilisation de la tomographie par émission de positrons (TEP). La stimulation électroacupuncturale (fréquence de 4 Hz/60 Hz en alternance de 2 secondes est appliquée sur le 36ES et le 4GI pendant 25 mn avant de mettre la queue de l’animal dans l’eau chaude à 47°C. Le temps de latence du retrait est mesuré. L’imagerie par TEP permet de visualiser les aires cérébrales impliquées qui correspondent à celles impliquées dans la douleur, mais inactivées par l’EA : le thalamus (zone postéro-inférieure du chiasma optique très proche de l’hypothalamus), une portion de l’insula et le gyrus cingulaire [[47]]. Le tableau IV récapitule les différentes aires cérébrales impliquées par l’EA.
Tableau IV. Principales structures du SNC observées par imagerie ou acupuncture expérimentale lors de l’EA analgésique.
Rhombencéphale (pont et bulbe rachidien) Formation réticulée Noyau raphé du pont Noyau raphé magnus Locus coeruleus Système limbique Gyrus cingulaire Amygdale Hippocampe Insula Hypothalamus
Conclusion
Les travaux de physiologie et d’imagerie concernant l’électroacupuncture dans les algies donnent une idée de plus en plus précise de la nature de ses mécanismes neurophysiologiques. Les données récentes suggèrent fortement l’implication de la libération des neuropeptides opioïdes (endorphines, enképhalines, dynorphine etc..), mais aussi l’action inhibitrice des récepteurs ionotropiques glutaminiques (en particulier NMDA), sans oublier la modulation de la douleur par le système inhibiteur descendant sérotoninergique et catécholaminergique. L’EA semble donc moduler le message nociceptif par de multiples mécanismes tout le long de sa transmission. L’imagerie démontre de manière formelle que les systèmes limbique, hypothalamique et le tronc cérébral sont activés par l’EA. Nous verrons dans un prochain article les modalités techniques de l’électroacupuncture et ses implications pratiques.
Références
[1]. Stéphan JM. Acupuncture expérimentale, stress et molécules informationnelles. Acupuncture & Moxibustion. 2006;5(2):162-170.
[2]. Stéphan JM. Acupuncture, récepteurs transmembranaires à tyrosine-kinases, à cytokines et transduction. Acupuncture & Moxibustion. 2007 Mars;6(1):79-86.
[3]. Zadina JE. Isolation and distribution of endomorphins in the central nervous system. Jpn J Pharmacol. 2002 Jul;89(3):203-8.
[4]. Stéphan JM. Acupuncture, récepteurs couplés aux protéines G et transduction. Acupuncture & Moxibustion. In press 2008.
[5]. Research Group of Acupuncture Anesthesia, P.M.C. The effect of acupuncture on the human skin pain threshold. Chin Med J. 1973;3:151-157.
[6]. Research Group of Acupuncture Anesthesia, P.M.C. The role of some neurotransmitters of brain in finger-acupuncture analgesia. Scientia Sinica. 1974;117:112-130.
[7]. Pomeranz B, Chiu D. Naloxone blockade of acupuncture analgesia : endorphin implicated.Life Sci.1976;19(11):1757-62.
[8]. Sjolund B, Terenius L, Eriksson M. Increased cerebrospinal fluid levels of endorphins after electro-acupuncture. Acta Physiol Scand. 1977;100(3):382-4.
[9]. Mayer DJ, Price DD, Rafii A. Antagonism of acupuncture analgesia in man by the narcotic antagonist naloxone. Brain Res.1977;121(2):368-72.
[10]. Clement-Jones V, McLoughlin L, Tomlin S, Besser GM, Rees LH, Wen HL. Increased beta-endorphin but not met-enkephalin levels in human cerebrospinal fluid after acupuncture for recurrent pain. Lancet. 1980;2(8201):946-9.
[11]. Chen XH, Han JS. Analgesia induced by electroacupuncture of different frequencies is mediated by different types of opioid receptors: another cross-tolerance study. Behav Brain Res. 1992;47(2):143-9.
[12]. Chen XH, Han JS. All three types of opioid receptors in the spinal cord are important for 2/15 Hz electroacupuncture analgesia. Eur J Pharmacol. 1992;211(2):203-10.
[13]. Han JS. Acupuncture and endorphins. Neurosci Lett. 2004;361(1-3):258-61.
[14]. Pan B, Castro-Lopes JM, Coimbra A. C-fos expression in the hypothalamo-pituitary system induced by electroacupuncture or noxious stimulation. Neuroreport. 1994;5(13):1649-52.
[15]. Pan B, Castro-Lopes JM, Coimbra A. Activation of anterior lobe corticotrophs by electroacupuncture or noxious stimulation in the anaesthetized rat, as shown by colocalization of Fos protein with ACTH and beta-endorphin and increased hormone release. Brain Res Bull. 1996;40(3):175-82.
[16]. Pan B, Castro-Lopes JM, Coimbra A. Chemical sensory deafferentation abolishes hypothalamic pituitary activation induced by noxious stimulation or electroacupuncture but only decreases that caused by immobilization stress. A c-fos study. Neuroscience. 1997;78(4):1059-68.
[17]. Yang J, Yang Y, Chen JM, Liu WY, Lin BC. Effect of hypothalamic supraoptic nucleus on acupuncture analgesia in the rat. Brain Res Bull. 2008;75(5):681-6.
[18]. Xie GX, Han JS, Hollt V. Electroacupuncture analgesia blocked by microinjection of anti-beta-endorphin antiserum into periaqueductal gray of the rabbit. Int J Neurosci. 1983;18:287-292.
[19]. Chao DM, Shen LL, Tjen-Alooi S, Pitsillides KF, Li P, Longhurst JC. Naloxone reverses inhibitory effect of electroacupuncture on sympathetic cardiovascular reflex responses. Am J Physiol Heart Circ Physiol. 1999;276:H2127-H2134.
[20]. Pomeranz B, Bibic L. Naltrexone, an opiate antagonist, prevents but does not reverse the analgesia produced by electroacupuncture. Brain Res.1988;452:227-231.
[22]. Han JS. Neurochemical basis of acupuncture. Annu Rev Pharmacol Toxicol.1982;22:193-220.
[23]. Jin WQ, Zhou ZF, Han JS. Electroacupuncture and morphine analgesia potentiated by bestatin and thiorphan administered to the nucleus accumbens of the rabbit. Brain Res. 1986;380:317-324.
[24].Yu LC, Han JS. Involvement of arcuate nucleus of hypothalamus in the descending pathway from nucleus accumbens to periaqueductal gray subserving an antinociceptive effect. Int J Neurosci. 1989;48:71-78.
[25].Wang Q, Mao L, Han JS. The arcuate nucleus of hypothalamus mediates low but not high frequency electroacupuncture in rats. Brain Res. 1990;513:60-66.
[26]. Takeshige C. The acupuncture point and its connecting central pathway for producing acupuncture analgesia. Brain Res Bull. 1993;30:53-67.
[27]. Chang Ht. Integrative action of thalamus in the process of acupuncture for analgesia. Scientia Sinica. 1973;16:25-60.
[28]. Gao M, Wang M, Li K, He L. Changes of mu opioid receptor binding sites in rat brain following electroacupuncture. Acupunct Electrother Res. 1997;22(3-4):161-6.
[29]. Tsai HY, Lin JG, Inoki R.Further evidence for possible analgesic mechanism of electroacupuncture:effects on neuropeptides and serotonergic neurons in rat spinal cord.Jpn J Pharmacol.1989;49(2):181-5.
[30]. Takeshige C, Sato T, Mera T, Hisamitsu T, Fang J. Descending pain inhibitory system involved in acupuncture analgesia.Brain Res Bull.1992;29(5):617-34.
[31]. Scherder EJ, Bouma A.Possible role of the nucleus raphe dorsalis in analgesia by peripheral stimulation: theoretical considerations.Acupunct Electrother Res. 1993;18(3-4):195-205.
[32]. Chang YH, Hiseh MT, Wu CR.Effects of acupuncture at pai-hui on the deficit of memory storage in rats.Am J Chin Med.1999;27(3-4):289-98.
[33]. Takagi J, Yonehara N. Serotonin receptor subtypes involved in modulation of electrical acupuncture. Jpn J Pharmacol. 1998;78(4):511-4.
[34]. Lee CK, Bae HS, Rho SW, Choi GS, Ko EJ, Kim SK, Lee YS, Lee HJ, Hong MC, Shin MK, Min BI, Kee HJ Molecular Changes in Remote Tissues induced by Electro-Acupuncture Stimulation at Acupoint ST36. Mol Cells. 2008;25(2).
[35]. Li A, Wang Y, Xin J, Lao L, Ren K, Berman BM, et al. Electroacupuncture suppresses hyperalgesia and spinal Fos expression by activating the descending inhibitory system. Brain Res. 2007;1186:171-9.
[36]. Koo ST, Lim KS, Chung K, Ju H, Chung JM. Electroacupuncture-induced analgesia in a rat model of ankle sprain pain is mediated by spinal alpha-adrenoceptors. Pain. 2008;135(1-2):11-9.
[37]. Kim HW, Uh DK, Yoon SY, Roh DH, Kwon YB, Han HJ, et al. Low-frequency electroacupuncture suppresses carrageenan-induced paw inflammation in mice via sympathetic post-ganglionic neurons, while high-frequency EA suppression is mediated by the sympathoadrenal medullary axis. Brain Res Bull. 2008;75(5):698-705.
[38]. Choi BT, Lee JH, Wan Y, Han JS. Involvement of ionotropic glutamate receptors in low frequency electroacupuncture analgesia in rats. Neurosci Lett. 2005 Apr 4;377(3):185-8.
[39]. Choi BT, Kang J, Jo UB. Effects of electroacupuncture with different frequencies on spinal ionotropic glutamate receptor expression in complete Freund’s adjuvant-injected rat. Acta Histochem. 2005;107(1):67-76.
[40]. Wang L, Zhang Y, Dai J, Yang J, Gang S. Electroacupuncture (EA) modulates the expression of NMDA receptors in primary sensory neurons in relation to hyperalgesia in rats. Brain Res. 2006;1120(1):46-53.
[41]. Xing GG, Liu FY, Qu XX, Han JS, Wan Y. Long-term synaptic plasticity in the spinal dorsal horn and its modulation by electroacupuncture in rats with neuropathic pain. Exp Neurol. 2007;208(2):323-32.
[42]. Ryu JW, Lee JH, Choi YH, Lee YT, Choi BT. Effects of protein phosphatase inhibitors on the phosphorylation of spinal cord N-methyl-d-aspartate receptors following electroacupuncture stimulation in rats. Brain Res Bull. 2008;75(5):687-91.
[43]. Wu MT, Sheen JM, Chuang KH, Yang P, Chin SL, Tsai CY, Chen CJ, Liao JR, Lai PH, Chu KA, Pan HB, Yang CF. Neuronal specificity of acupuncture response : a fMRI study with electroacupuncture. Neuroimage. 2002;16(4):1028-37.
[44]. Zhang WT, Jin Z, Huang J, Zhang L, Zeng YW, Luo F, Chen AC, Han JS. Modulation of cold pain in human brain by electric acupoint stimulation : evidence from fMRI. Neuroreport. 2003;14(12):1591-6.
[45]. Zhang WT, Jin Z, Cui GH, Zhang KL, Zhang L, Zeng YW, Luo F, Chen AC, Han JS. Relations between brain network activation and analgesic effect induced by low vs. high frequency electrical acupoint stimulation in different subjects: a functional magnetic resonance imaging study. Brain Res. 2003;982(2):168-78.
[46]. Napadow V, Makris N, Liu J, Kettner NW, Kwong KK, Hui KK. Effects of electroacupuncture versus manual acupuncture on the human brain as measured by fMRI. Hum Brain Mapp. 2005;24(3):193-205.
[47]. Maenaka T, Tano K, Nakanishi S, Tsukada H, Ishida T. Positron emission tomography analysis of the analgesic effects of acupuncture in rhesus monkeys. Am J Chin Med. 2006;34(5):787-801.
Parlement Hongrois (néo-gothique -éclectique) 1885-1904 – Budapest – Hongrie
Résumé. Introduction. En France en 2016, la prévalence globale du diabète était estimée à 5% de la population, à laquelle s’ajoute celle de près de 20% de syndrome métabolique responsable à plus ou moins long terme de ce même diabète de type 2 et des complications cardio-vasculaires. L’acupuncture et l’électroacupuncture (EA) peuvent-elles avoir leur place dans la thérapeutique classique ? Méthodes. A partir d’une étude de cas d’un homme de 51 ans présentant un diabète de type 2 avec traitement usuel, un protocole agissant selon la différenciation des syndromes (bianzheng) sur le Vide de yin et de yang des Reins a été appliqué par EA à la fréquence alternée 3/80 Hz (durée de l’impulsion : 300µs) sur 6MC (neiguan), 6Rt (sanyinjiao), une EA à 15Hz (200µs) sur 36E (zusanli), 12VC (zhongwan) et 4VC (guanyuan) ; et enfin une puncture manuelle avec recherche du deqi sur les points 23V (shenshu), 3R (taixi), 4Rt (gongsun), 7C (shenmen), 20VG (baihui), yishu (EX-B3). Le critère principal de jugement est un test de glycémie aléatoire effectué avant et après la séance, quinze séances au total sur une période de dix-neuf mois. Le critère secondaire de jugement est l’effet à long terme mesuré par l’évolution de l’hémoglobine glyquée (HbA1c). Résultats. Après une seule séance d’acupuncture et EA, on observe une baisse statistiquement significative de la glycémie (DM=0,498mm/L ; IC 95% ; 0,2035 à 0,7925 ; p=0,0027, test de Student) entre les deux groupes de mesure et une tendance à la baisse globale de la HbAC1. Conclusion. L’acupuncture et EA doivent être intégrées au panel de soins usuel dans le cadre de la médecine intégrative. L’effet est immédiat et dure dans le temps. Mots clés. Diabète de type 2 – acupuncture – électroacupuncture – effets immédiats – insulinorésistance – bianzheng.
About a clinical case of type 2 diabetes: immediate effects of acupuncture and electroacupuncture. What are the long-term effects?
Summary.Introduction. In France in 2016, the overall prevalence of diabetes was estimated at 5% of the population, to which should be added the prevalence of nearly 20% of metabolic syndrome responsible in the more or less long term for type 2 diabetes and cardiovascular complications. Can acupuncture and electroacupuncture (EA) have a place in conventional therapy? Methods. Based on a case study of a 51 year old man with type 2 diabetes with standard treatment, a protocol acting according to the differentiation of syndromes (bianzheng) on the yin and yang empty of the Kidneys was applied by EA at the alternating frequency of 3/80 Hz (pulse duration: 1.5 seconds): 300µs) on 6P (neiguan), 6SP (sanyinjiao), an EA at 15Hz (200µs) on 36ST (zusanli), 12REN (zhongwan) and 4REN (guanyuan) ; and finally a manual puncture with deqi search on 23BL (shenshu), 3KID (taixi), 4SP (gongsun), 7HE (shenmen), 20DU (baihui), yishu (EX-B3). The main outcome is a random blood glucose test carried out before and after the session, a total of 15 sessions over a period of 19 months. The secondary outcome is the long-term effect measured by the change in glycated hemoglobin (HbA1c). Results. After a single acupuncture session and EA, there was a statistically significant decrease in blood glucose (DM=0.498mm/L; 95% CI; 0.2035 to 0.7925; p=0.0027, Student’s test) between the two measurement groups and an overall downward trend in HbAC1. Conclusion. Acupuncture and EA should be integrated into the usual care panel in integrative medicine. The effect is immediate and lasts over time. Keywords. Diabetes type 2 – acupuncture – electroacupuncture – immediate effects – insulin resistance – bianzheng.
Introduction
En France, la prévalence globale du diabète était estimée à 5% de la population en 2016 dont 90% des cas correspondaient au diabète de type 2 (DT2), chiffre d’ailleurs largement sous-estimé puisqu’il ne tient pas compte des personnes non traitées ou non diagnostiquées. Or, compte tenu du caractère silencieux de la maladie, on estime que de 20 à 30% des adultes diabétiques ne sont pas diagnostiqués. C’est également 6% de la mortalité mondiale et 50% des décès liés au diabète sont dus à une maladie cardiovasculaire.
L’incidence du DT2 augmente avec l’âge. La maladie se manifeste généralement après 40 ans et elle est diagnostiquée à un âge moyen proche de 65 ans. L’incidence est maximale entre 75 et 79 ans avec 20% des hommes et 14% des femmes traités pour cette maladie. La prévalence de la maladie a beaucoup augmenté ces dernières années. Ceci est essentiellement lié au vieillissement de la population, mais aussi aux déséquilibres nutritionnels, à la sédentarité qui engendrent une hygiène de vie délétère, induisant une augmentation générale du poids et du nombre de personnes obèses. Un autre sujet d’alerte est l’augmentation récente de diabètes iatrogènes, associés en particulier à certains neuroleptiques [[1]].
Définition
Le diabète de type 2 est défini par :
– une glycémie supérieure à 1,26 g/l (7,0 mmol/l) après un jeûne de 8 à 12 heures et vérifiée à deux reprises,
– ou la présence de symptômes de diabète (polyurie, polydipsie, amaigrissement) associée à une glycémie (sur plasma veineux) supérieure ou égale à 2 g/l (11,1 mmol/l),
– ou une glycémie (sur plasma veineux) supérieure ou égale à 2 g/l (11,1 mmol/l) deux heures après une charge orale de 75 g de glucose (critères proposés par l’Organisation Mondiale de la Santé).
Le diabète de type 2 accélère l’athérosclérose et provoque des complications microvasculaires (rétinopathie, néphropathie et neuropathie) et macrovasculaires (infarctus du myocarde, artérite et accident vasculaire cérébral). L’objectif du traitement du patient atteint d’un diabète de type 2 est de réduire la morbi-mortalité, par l’intermédiaire notamment d’un contrôle glycémique correct et d’obtenir une hémoglobine glyquée (HbA1c) inférieure ou égale à 7% [[2]].
Le traitement
Les mesures hygiénodiététiques et l’activité physique sont toujours nécessaires, et doivent être réévaluées à chaque étape de la stratégie. Un amaigrissement, même limité (- 5 % du poids), apporterait un bénéfice glycémique démontré en cas d’excès pondéral. Une activité physique modérée de 2 h 30 par semaine doit être mise en place progressivement. La monothérapie avec la metformine est le traitement oral de première intention. Ce n’est qu’en cas d’intolérance ou de contre-indication à la metformine que l’on peut choisir entre de multiples possibilités thérapeutiques comme :
– les sulfamides hypoglycémiants et les glinides qui stimulent la production d’insuline au niveau du pancréas ;
– les inhibiteurs des alpha-glucosidases (par exemple acarbose) qui retardent l’absorption des glucides après les repas ;
– les agonistes du récepteur du glucagon-like peptide-1 (GLP1) qui ralentissent la vidange gastrique, limitent l’appétit et stimulent la sécrétion d’insuline, mais uniquement en cas d’élévation de la glycémie, limitant donc le risque d’hypoglycémie. Ils peuvent d’ailleurs être combinés à une insuline dite « lente » qui maintient continuellement une concentration basale d’insuline ;
– les inhibiteurs de la dipeptidylpeptidase 4 (DPP4) ou gliptines qui bloquent la dégradation du GLP1 ;
– les inhibiteurs du co-transporteur sodium/glucose de type 2 (SGLT2) comme la dapagliflozine, qui agissent sur le rein en bloquant la réabsorption du glucose.
La recommandation de la Haute Autorité Française évoque la possibilité de passer rapidement à l’insuline si l’écart à l’objectif glycémique est supérieur à 1%, ce qui survient souvent après plusieurs années d’évolution, d’où le terme de diabète insulino-requérant. On comprendra ainsi que, l’ancien terme diabète non insulinodépendant (DNID) ait été abandonné pour diabète de type 2, du fait d’une insulinopénie progressive et inéluctable.
Les variations glycémiques
Les variations glycémiques observées chez le sujet non diabétique sont surtout en rapport à la réponse métabolique post-prandiale liée aux repas. C’est physiologique et reproductible d’un jour à l’autre dans la mesure où les activités et les repas sont semblables. Classiquement, après la prise d’un repas, la glycémie s’élève dans les 15 minutes suivant l’absorption orale pour atteindre un pic aux alentours de la 45e minute. En règle générale, on a un retour au niveau basal de la glycémie dans l’heure qui suit le pic glycémique post-prandial. Puis le niveau glycémique reste alors stable jusqu’au prochain repas chez le patient non diabétique.
Chez le patient diabétique sous insuline, cette variabilité glycémique est en revanche directement liée à la perte de l’homéostasie glycémique en rapport avec la diminution puis la carence de la sécrétion endogène insulinique [[3]].
De ce fait, il paraît intéressant d’observer si l’acupuncture et techniques associées, en l’occurrence ici l’électroacupuncture (EA) permet d’agir sur cette variabilité glycémique. Ainsi, l’objectif de cette étude, évaluant un seul cas clinique, mais observé sur dix-neuf mois, est de constater les effets immédiats du traitement acupunctural, mais aussi d’observer si on peut maintenir un diabète équilibré sur un long terme.
Matériel et Méthodes
Observation
Monsieur D. E., 51 ans présente un diabète de type 2 devenu requérant, associé à une hypertension artérielle (HTA) et un état dépressif depuis 2008, ayant nécessité à partir de 2015 sa mise en arrêt de travail longue durée, puis sa mise en invalidité. Non sportif, il pèse 80kgs pour 1m65 et son indice de masse corporelle est de 29,4 objectivant un surpoids, à la limite de l’obésité modérée [note 1]. Le dosage sanguin de l’hémoglobine glyquée (HbA1c) est à 8% quand il est vu pour la première fois en 2013. Le traitement anti-diabétique associe metformine 500 mg, exénatide 5µg (analogue du GLP1), insuline glargine : 120 unités le soir en moyenne à adapter en fonction de la glycémie ; irbesartan 300 mg, manidipine 20mg pour l’HTA ; atorvastatine en raison d’une hypercholestérolémie et enfin miansérine 60 mg pour son état dépressif. La dose de metformine n’a pu être augmentée en raison des nombreux troubles gastro-intestinaux.
Au départ la langue est sèche et rouge ; le pouls est profond(chen), fin(xi) et rapide (shuo) qui correspond selon la différenciation des syndromes (bianzheng) à un Vide de yin des Reins.
Traitement acupunctural
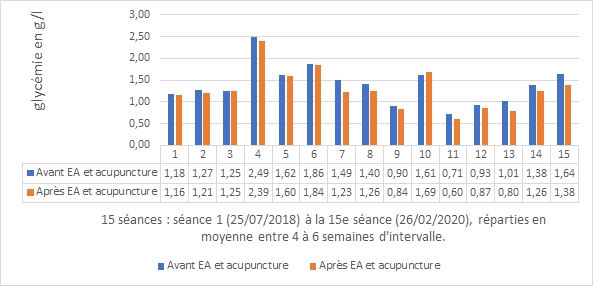
Le traitement est individualisé à partir de 2013 mettant l’accent tout d’abord sur ce Vide de yin des Reins, comme cela a déjà été décrit par exemple par Hawawini [[4]]. Mais progressivement, au cours des mois de traitement, il y a évolution vers un Vide de yin et de yang des Reins, conséquence de ce Vide de yin des Reins avec une langue devenant pâle, un pouls toujours profond(chen) et fin(xi). En accord avec le patient, un protocole de traitement est mis en place au cabinet médical lors de la séance du 25 juillet 2018 et poursuivi sur dix-neuf mois à raison d’une séance de 20 à 25 minutes réalisée entre quatre et huit semaines d’intervalle, soit quinze séances jusqu’au 26 février 2020.
Un test de glycémie aléatoire [note 2], par mesure de la glycémie capillaire est effectuée par le patient lui-même, avant et après la séance avec un lecteur Freestyle Optium Abbot®. Le prélèvement de la goutte de sang s’effectue par une lancette à usage unique pour chaque contrôle par piqûre sur le côté de la dernière phalange d’un des trois derniers doigts. La séance est généralement réalisée chaque mois à la même heure : 17h. Le critère de jugement principal est donc d’objectiver l’effet immédiat de la séance d’acupuncture, à savoir une diminution de la glycémie après la séance. Le critère secondaire est l’effet à long terme mesuré par l’évolution de l’hémoglobine glyquée (HbA1c).
Analyse statistique
Il s’agit d’une étude de cas réalisée sur un seul patient avec mesure de la glycémie avant, puis après quinze séances d’acupuncture et EA, réparties à une moyenne d’un mois d’intervalle sur une durée de dix-neuf mois du 25/07/2018 au 26/02/2020. Il a été estimé que la taille des mesures serait suffisante pour fournir des résultats qui répondraient à notre critère de jugement principal. Le test t de Student pour échantillons appariés a été choisi. Ce test t permet de comparer deux mesures d’une variable quantitative effectuées sur le même sujet. En fait, ce test traite les deux échantillons appariés comme un seul sur lequel on aurait mesuré la différence entre les deux mesures. On compare donc les moyennes de deux échantillons appariés. Les hypothèses à tester sont : H0 hypothèse nulle = les moyennes des deux groupes sont identiques : μ1 = μ2 ; HA = les moyennes des deux groupes sont différentes : μ1 ≠ μ2. Après calcul du test t, la probabilité d’obtenir cette valeur t est calculée de façon à savoir si l’H0 est vraie (c’est-à-dire si les moyennes dans les populations sont égales). La différence statistiquement significative est fixée à p=0,05. Si cette probabilité p calculée est inférieure à p<0,05, alors, on ne peut pas accepter H0 et on conclut dans ce cas à la validité de l’hypothèse HA, à savoir que les glycémies diffèrent.
Les analyses ont été effectuées en utilisant le logiciel statistique biostaTGV [[5]].
Résultats
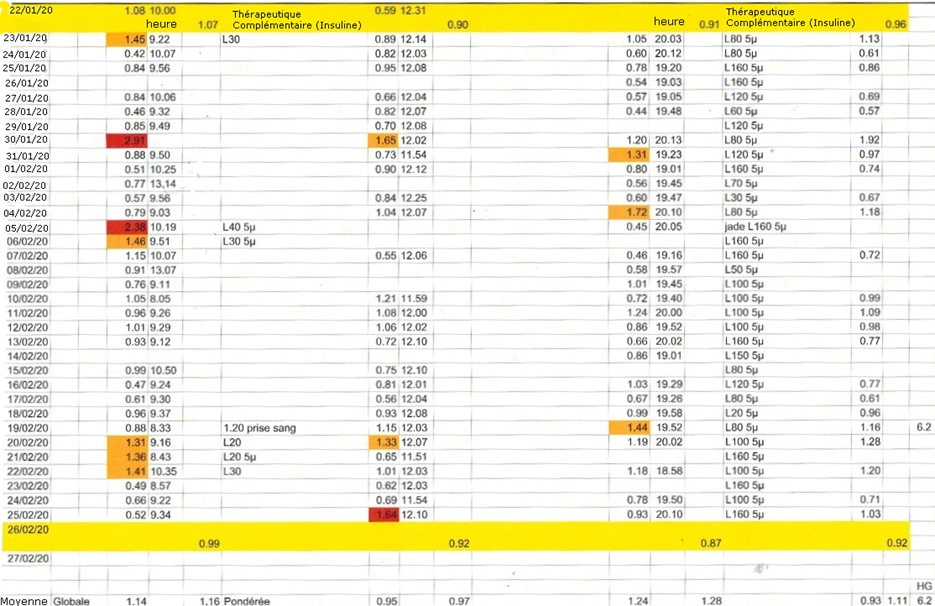
Le tableau I objective les dates des séances et les glycémies avant et après les quinze traitements. Sont notifiés également les dosages de la HBA1c.
Résultats du test statistique à 14 degrés de liberté observée t = 3,6266 ; p=0,00274 ; en valeur absolue différence moyenne (DM = 0,498mm/L avec un intervalle de confiance à 95% [0,2035 à 0,7925] entre les deux groupes de mesure.
En conclusion, l’hypothèse HA est validée objectivant une diminution statistiquement significative (p=0,0027, test de Student) des taux glycémiques en mm/l avant et après séance.
Les résultats sont présentés dans le tableau 1, les figures 1 et 2.
Le critère secondaire mesuré objective une baisse globale de la HbAC1. Ainsi à la toute première séance en 2013, l’HbA1c était à 8% alors qu’à la dernière en février 2020, le chiffre était à 6,2% (tableau I). On observe donc une baisse continue de son taux d’hémoglobine glyquée sur une période de sept ans, permettant de ne pas augmenter la thérapeutique usuelle.
Tableau I. Dates des séances. Glycémie avant et après séance en mmol/L. Dosage de l’hémoglobine glyquée HbA1c en %.
Dates
avant
après
différence
HbA1c
25/07/2018
6,49
6,38
0,11
7,1
18/09/2018
6,98
6,65
0,33
14/11/2018
6,87
6,87
0
6,9
26/12/2018
13,70
13,14
0,56
29/01/2019
8,91
8,80
0,11
26/02/2019
10,23
10,12
0,11
09/04/2019
8,19
6,76
1,43
15/05/2019
7,70
6,93
0,77
19/06/2019
4,95
4,62
0,33
6,8
28/08/2019
8,85
9,30
-0,45
23/10/2019
3,90
3,30
0,6
19/11/2019
5,11
4,78
0,33
17/12/2019
5,55
4,40
1,15
22/01/2020
7,59
6,93
0,66
26/02/2020
9,02
7,59
1,43
6,2
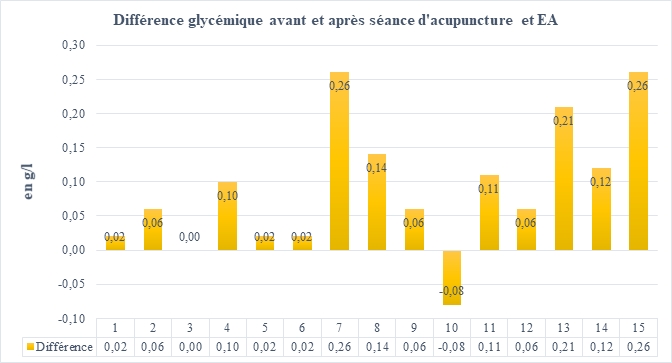
Figure 1. Mesure de la glycémie en g/L avant et après une séance d’acupuncture et EA pour le patient et évaluée quinze fois.
Figure 2. Différences entre la glycémie avant et après la séance d’acupuncture et d’électroacupuncture.
Discussion
L’objectif de cette étude était de démontrer que l’acupuncture associée à l’électroacupuncture a un effet immédiat sur les paramètres métaboliques. A été analysée ainsi la glycémie avant et après une séance de vingt minutes. On observe donc une diminution statistiquement significative de cette glycémie (p=0,00269).
On notera aussi que les glycémies effectuées à domicile sont généralement dans les normes. Les doses d’insuline glargine et d’exénatide sont modulées en fonction de la glycémie. On observe quelques pics, mais globalement Monsieur D. E. est stabilisé tout comme le montre son HbA1c (Tableau II). Il semble donc que l’acupuncture et techniques associées aient un effet favorable sur le diabète de type 2 requérant.
Les études expérimentales en acupuncture montrent-elles également cet effet ? Peut-on corréler cette étude de cas clinique à des essais comparatifs randomisés ou mieux des méta-analyses ? La discussion va permettre d’approfondir ces problématiques.
Tableau II. Glycémies à jeun réalisées à domicile trois fois par jour en moyenne, vers 9h30, midi et en début de soirée durant la période du 22 janvier au 26 février 2020. Dose d’insuline glargine (L30 en Unités) et d’exénatide (en µg).
Etudes expérimentales
En effet, des études sur l’animal suggèrent cet effet favorable.
Notons que les études ont été faites à la fois sur des modèles de diabète type 1 comme les rats diabétiques induits par la streptozotocine (STZ) [note 3] ou des modèles de type 2 comme le rat des sables (Psammomys obesus) [note 4], le rat GK (Goto-Kkizaki) japonais dont le diabète est consécutif à une alimentation relativement hypercalorique avec une obésité modérée, ou autres modèles animaux de diabète induit par un régime alimentaire hypercalorique ou rats en résistance à l’insuline induite par injection de stéroïdes (SIIR).
Effets immédiats de l’EA
Tous les travaux expérimentaux suggèrent une action immédiate de l’EA à 30, 60 ou 90 mn engendrant une baisse rapide de la glycémie versus groupes placebo ou témoin. La régulation de la glycémie se fait par augmentation de la sécrétion de β endorphines [6-8], de l’insuline ou de sa sensibilité [6,7,9-11] ; par la diminution de la résistance à l’insuline, soit du fait de la baisse des acides gras plasmatiques [11-14], soit par régulation à la hausse de l’expression des protéines de signalisation de l’insuline (IRS1, GLUT4 ou Akt2) [11-15]. Autres actions de l’acupuncture et de l’EA sur la baisse immédiate de la glycémie : stimulation des nerfs cholinergiques [13,15,16] et par augmentation de la nNOS [13].
On notera que, dans la grande majorité des cas, la fréquence appliquée sur points (souvent 36E zusanli, 12VC zhongwan et 4VC guanyuan [[17]])est une fréquence à 15Hz pour une durée d’impulsion de 200µs (voir annexe 2 pour de plus amples détails : Expérimentation animales sur les effets immédiats de l’acupuncture et électroacupuncture), d’où le choix de la fréquence utilisée dans ce cas clinique pour obtenir un effet immédiat. Outre ces effets, on a pu objectiver que ces effets peuvent se prolonger comme le démontrent d’autres travaux expérimentaux.
Effets à long terme
Shapira et coll. en 2000 objectivaient que l’électroacupuncture (15Hz ; 350 µs) appliquée au zhongwan (12VC), gwanyuan (4VC) induisait un effet hypoglycémiant prolongé sur un modèle de rats diabétiques de type 2 (rats Psammomys) versus l’électroacupuncture placebo. Cet effet se prolonge sur plus de deux semaines après la fin de l’intervention de manière statistiquement significative (p<0,0001) versus groupes placebo et témoin, sans qu’un effet hypoinsulinémique ne soit mis en évidence [[18]]. Mêmes observations chez les rats diabétiques Zucker obèses, l’EA (10 Hz, largeur de l’impulsion 200 µs, 30 mn 3 fois par semaine) sur zhongwan (12VC) et gwanyuan (4VC) engendre une baisse significative de la glycémie 30 mn après la séance aux 5e, 8e et 12e jour de traitement, plus importante que l’EA au zusanli (36E) [[19]].
Comme on l’a vu dans les effets immédiats, l’EA a donc également un effet hypoglycémiant important sur les effets à long terme. L’acupuncture et EA agiraient sur l’insulino-résistance, en améliorant la fonction des cellules bêtas des îlots de Langherans et en allégeant le dysfonctionnement endothélial [[20]].
Les études expérimentales ont objectivé diverses voies potentielles concernant l’action de l’acupuncture et EA sur l’insulino-résistance.
Les mécanismes s’effectuent ainsi par régulation de l’hyperactivité de l’axe hypothalamo-hypophyso-surrénalien (réduction de la corticotropin-releasing hormone (CRH) et du cortisol) et de ce fait action sur le métabolisme des lipides sanguins (triglycérides, cholestérol HDL et LDL) [[21]] ; par régulation à la hausse des concentrations de certaines protéines des tissus cibles de l’insuline (foie et muscle squelettique), tels que la protéine kinase activée par l’AMP (AMPK) [22-24], les protéines de signalisation de l’insuline (IRS1, GLUT4 ou Pi3K/Akt) [[25]], la protéine SREBP-1 [[26]], par régulation de l’expression des protéines hypothalamiques phosphatidylinositol-3-kinase (PI3K)-p85 [[27]] chez les rats obèses, par régulation des protéines de synthèse des acides gras [23,24,26], en supprimant la réaction inflammatoire du tissu hépatique et en régulant l’adiponectine et la leptine sérique [24,[28]] ; en réduisant les niveaux des espèces réactives d’oxygène (ROS) et le contenu de malondialdéhyde (MDA) [[29]] ; en atténuant les lésions structurelles de l’îlot du pancréas et l’apoptose des cellules β pancréatiques chez les rats diabétiques [34] ; et enfin, par une action épigénétique [note 5] [30-34]. En conclusion, de multiples niveaux et divers systèmes sont impliqués dans les actions de l’acupuncture et EA dans l’amélioration de l’insulino-résistance.
Ces différentes études confortent donc dans ce cas clinique le choix des points en EA et de la fréquence appliquée (3/80Hz ; 300µs [note 6] sur 6MC (neiguan), 6Rt (sanyinjiao) (voir annexe 3 : Expérimentation animales sur les effets à long terme de l’acupuncture et électroacupuncture), surtout qu’on observe également un effet à long terme sur la prévention des complications diabétiques et des maladies cardiovasculaires [25].
Néanmoins, le niveau de preuve scientifique soutenu par les études expérimentales n’est pas suffisant et il convient de se reporter aux essais comparatifs randomisés pour alléguer un grade de recommandation dans l’action de l’acupuncture et/ou EA dans le diabète de type 2.
Les essais comparatifs randomisés (ECR) et méta-analyses
Effets immédiats
Concernant les effets immédiats de l’acupuncture et/ou de l’EA, il n’existe aucune méta-analyse sur le sujet. Néanmoins quatre études cliniques pilotes existent [35-38].
Belivani et col. en 2015 [35] dans leur étude pilote pragmatique randomisée ont comparé trois groupes de femmes en obésité (IMC> 30 kg/m²). Le critère de jugement était la glycémie mesurée avant et après l’administration en une seule séance (30 min) d’EA 2/80Hz (durée impulsion100µs) [note 7]. Trois groupes : un groupe Dorsal (n=5) avec EA sur les points ganshu (18V), pishu (20V), weishu (21V) et shenshu (23V) bilatéralement (correspondant aux niveaux segmentaires de l’innervation du pancréas) ; le groupe Oreille (n=5) avec EA aux points d’oreille Poumon et Cœur dans la cavum conchae ; le groupe Corps (n=6) avec EA sur shousanli (10GI), quchi (11GI), zusanli (36E) et zhongping [note 8]. Ils observaient après la séance une diminution significative de la glycémie dans les groupes Dorsal et Corps alors qu’il n’y avait aucun changement et même une tendance vers une hyperglycémie dans le groupe Oreille.
Mohanty et coll. en 2016 [36] se sont aussi intéressés aux effets immédiats de l’acupuncture sur la glycémie de trente-six sujets sains hommes et femmes âgés de 18 à 24 ans. Il s’agit d’un ECR contre placebo et en double aveugle (participants et investigateur). Deux groupes : acupuncture manuelle au zhongwan 12VC (n=18) et groupe acupuncture témoin placebo (n=18) sur un non-point à 1 cun latéral au 12VC. Le critère de jugement est la glycémie aléatoire mesurée avant et après une seule séance (20 min, faite entre 10h30 et 11h30. Ils observent une tendance à la diminution de la glycémie chez les sujets sains mais non statistiquement significative dans les groupes acu et témoin avant et après séance et entre les deux groupes.
Ce qui se rapproche davantage du cas clinique présenté ici est l’ECR contre placebo et en double aveugle également (participants et investigateur) chez quarante diabétiques hommes de type 2 (âge moyen 56±11 ans) [37]. Le protocole de cet ECR est identique à celui de Mohanty [36] avec le même point utilisé zhongwan 12VC (n=20) dans le groupe traité. Après une seule séance d’acupuncture, les auteurs observent une diminution statistiquement significative (p<0,001) de la glycémie dans le groupe acupuncture avant et après séance, alors qu’il n’y a pas de changements dans le groupe témoin.
On observe également une baisse de la glycémie après 30 mn d’acupunture sur zusanli (36E)avant etaprès une unique séance dans l’ECR de Mooventham et coll. dans le groupe Acu (n=30) versus groupe placebo (n=30) [38].
Seuls deux ECR objectivent donc une efficacité immédiate significative dans le diabète de type 2 et sur un seul point : acupuncture manuelle sur 12VC [37] et sur E36 [38]. On retrouve la même efficacité statistiquement significative avec l’EA (2/80 Hz ;100µs) mais chez les femmes obèses non diabétiques [34] alors qu’un autre ECR sur des sujets sains [36] stimulant 12VC, la baisse est légère et non significative. Cela montre l’action cybernétique de l’acupuncture : celle-ci n’agit que s’il y a rupture de l’homéostasie.
Effets à long terme
La première méta-analyse concernant acupuncture et diabète de type 2, au total vingt et un ECR (n=1943 participants) a été publiée en août 2019. Combinée ensemble aux traitements classiques, l’acupuncture manuelle versus acupuncture simulée ou sans, offre une réduction statistiquement significative de la glycémie (MD= 1,21mmol/l, IC à 95% : 1,56 à 0,87 ; I²=91%), glycémie postprandiale à 2h (MD : 2,13 mmol/l ; IC à 95% : 2,79 à 1,46), HA1c (MD= 1,12% ; IC à 95% = 1,62 à -0,62). Les auteurs montrent également en critères secondaires de jugement que l’acupuncture peut améliorer le taux des lipides sanguins, le contrôle de la pression artérielle et réduire le poids. Toutefois, la puissance de cette méta-analyse est faible avec de nombreux ECR de petite taille, une mauvaise qualité méthodologique des ECR examinés et donc la quantité de preuves n’est pas entièrement convaincante, surtout qu’il existe une grande hétérogénéité. Ainsi le test d’Higgins I² = 91% [note 9] peut s’expliquer entre autres par le fait que les auteurs ont analysé en même temps l’acupuncture manuelle de type Médecine Chinoise, l’auriculothérapie, l’acupression, l’enfouissement de catgut sur les acupoints, EA, etc.. Quoi qu’il en soit les auteurs concluent à la nécessité d’études bien planifiées, à long terme et de bonne qualité méthodologique [[39]].
La seconde méta-analyse de Wu et coll. s’est davantage axée sur le traitement de l’insulino-résistance (IR) [note 10]. Vingt études (n=1639) ont été analysées. Dans les 20 études incluses, les participants avaient reçu un diagnostic de maladies liées à l’IR, dont 8 (40 %) patients atteints de diabète de type 2, 8 (40%) avec un syndrome ovarien polykystique (SOPK), 2 (10%) avec le syndrome métabolique, 1(5%) avec hypertension essentielle et 1 (5%) avec une obésité abdominale. L’acupuncture et techniques associées seules ou combinées au traitement occidental ont été comparées versus acupuncture factice (3 ECR) ou versus traitements occidentaux seuls (17 ECR). Avec l’acupuncture, l’évaluation du modèle d’homéostasie de la résistance à l’insuline (Homeostasis Model Accessment of insuline resistance HOMA-IR) [note 11] a diminué de manière significative (différence moyenne (MD) = -1,04, intervalle de confiance à 95% (IC) -1,37 à -0,71 ; p<0,00001 ; I²=95%), tout comme la glycémie à jeun (MD=-0,56, IC à 95% -0,88 à -0,25; p=0,0005 ; I²=96%), la glycémie postprandiale à 2h (2hPG) (MD = -0,91, IC à 95%, -1,62 à -0,20 ; p=0,01 ; I²=96%) et l’insuline à jeun (MD = -3,23, IC à 95% -4,14 à -2 ; p<0,00001 ; I²=90%). On remarque en même temps que l’indice de sensibilité à l’insuline (ISI) (MD= 0,36, IC à 95% 0,18 à 0,53 ; p<0,0001 ; I²=84%) a augmenté et que surviennent moins d’événements indésirables [[40]]. Là encore et comme la précédente méta-analyse, tout ceci suggère que l’acupuncture peut avoir une valeur thérapeutique dans le traitement de la résistance à l’insuline. Toutefois, les conclusions sont limitées par des lacunes méthodologiques des ECR inclus et la grande hétérogénéité.
La méta-analyse similaire de Song et coll. (9 ECR, n=562) s’intéressant à l’efficacité de l’acupuncture sur la résistance à l’insuline et analysant les ECR jusqu’en janvier 2018 a objectivé également des effets significatifs concernant HOMA-IR (MD= 0,70, IC à 95% 0,04 à 1,35, p=0,04<0,05) et sur les résultats secondaires comme la glycémie à jeun, l’insuline à jeun et de la glycémie postprandiale à 2 h. Mais les auteurs signalaient que même si l’acupuncture pouvait aider à améliorer l’IR dans une certaine mesure, il était nécessaire de confirmer par d’autres ECR de haute qualité [[41]].
Une méta-analyse (5 ECR) étudiant la moxibustion dans le diabète de type 2 n’a pas objectivé de réelle efficacité, et il faudra davantage d’ECR de bonne qualité méthodologique pour affirmer une efficacité [[42]].
En revanche et plus récemment, la méta-analyse de Li et coll. parue en octobre 2020 s’est intéressée au syndrome métabolique [note 12]. Un total de 13 ECR ont été identifiés (n=834). Après quatre à douze semaines de traitement (de 20 à 72 séances d’acupuncture), les auteurs observent que l’acupuncture offre une diminution statistiquement significative par rapport à l’acupuncture simulée de divers indices (tour de taille, perte de poids avec réduction de l’indice de masse corporelle IMC, amélioration de l’HbA1c, des TG, du cholestérol total et de l’HTA) ; améliore également le contrôle des taux de triglycérides et d’HDL cholestérol versus traitement conventionnel. Par ailleurs, il est observé également que l’ajout de l’acupuncture au traitement conventionnel engendre des effets globaux nettement plus importants que le traitement conventionnel seul, en ce qui concerne la circonférence de la taille abdominale, l’IMC (DM=−2,54 (Z= 5,83, IC à 95%= -3,40 -1,69) ; p <0,001), ainsi que des améliorations sur la plupart des indices hyperlipidémiques et de la glycémie à jeun. L’acupuncture est donc bénéfique dans le traitement des syndromes métaboliques seule ou associée au traitement conventionnel. Néanmoins, encore une fois, en raison d’une grande hétérogénéité, des ECR à plus grande échelle sont encore nécessaires pour confirmer l’efficacité de l’acupuncture dans le syndrome métabolique [[43]].
En conclusion, selon les preuves fondées sur des données probantes, il est encore nécessaire de réaliser des ECR de grande puissance et de haute qualité méthodologique avant d’affirmer que l’acupuncture et techniques associées puissent être totalement efficaces sur un long terme dans le diabète de stade 2.
Le choix des points et de stimulation
Il existe un consensus dans le choix des points zusanli (36E) et zhongwan (12VC), points classiques utilisés à la fois dans les études expérimentales et les ECR. Feng et coll. en 2018 avaient montré que les points les plus utilisés dans le traitement du diabète de type 2 étaient par ordre de fréquence dans les ECR parus entre 2014 et 2018, 36E, 23V, 6Rt, yishu, 12VC, 11GI (quchi), 9Rt (yinlingquan), 6MC, 4GI, 10Rt (xuehai), 40E (fenglong) et 20V (guanyuan) [17].
On note que le point yishu (EX-B3) est situé dans la zone métamérique et segmentaire du viscérotome innervé par les nerfs cholinergiques du système parasympathique (8e vertèbre thoracique (T8) qui stimulent les sécrétions du pancréas exocrine, mais aussi celles du pancréas endocrine avec sécrétion d’insuline et inhibition du glucagon.
Autre point important à soulever : l’EA est-elle équivalente à l’acupuncture manuelle ?
En d’autres termes, l’acupuncture manuelle (AM) avec recherche du deqi a-t-elle les mêmes effets physiologiques que l’électroacupuncture ? Cette question pragmatique a été en partie analysée à partir des ECR. On s’est aperçu que les techniques de stimulation de l’aiguille en acupuncture manuelle sont presque toujours appliquées pour une durée beaucoup plus courte que la stimulation électrique (c.-à-d. en secondes plutôt qu’en minutes). D’autre part, souvent dans les ECR d’EA, la stimulation manuelle est fréquemment effectuée brièvement en premier afin d’obtenir le deqi, suivie ensuite d’EA, d’où un biais méthodologique dans le sens où l’AM est effectivement comparée à une combinaison d’AM plus EA. D’où l’existence de nombreux facteurs de confusion si la comparaison s’effectue entre AM versus EA, ou AM par rapport à AM+EA.
Autre biais : l’EA est typiquement appliquée au moins 15 à 20 minutes, alors que l’AM en stimulation en continu n’est jamais réalisée. En effet, 20 mn de stimulation manuelle serait trop algique et de ce fait, les aiguilles sont stimulées quelques secondes puis laissées en place 20 mn. La recherche comparative de l’efficacité de l’AM versus EA en 2015 par Langevin et coll. [[44]] a retrouvé un seul ECR comparant EA versus AM [[45]] et seize autres ECR comparant AM par rapport à AM combinée à l’EA. La plupart des ECR portait sur l’évaluation de l’efficacité analgésique (n=10 [59%]). Sept ECR ont objectivé dans cette indication que l’AM+EA avait un effet antalgique supérieur par rapport à AM seule. Une meilleure analgésie était retrouvée si l’EA prolongée est ajoutée à la stimulation manuelle comparée à une stimulation manuelle brève ou intermittente.
Sept méta-analyses comprenaient une comparaison quantitative des résultats entre AM et EA. Une seule concernant la gonarthrose a réalisé une comparaison directe entre AM et EA et a objectivé que quatre ECR (n=614) montraient que l’EA était supérieure à l’AM seule (p=0,042) [[46]].
Les autres comparaisons indirectes entre EA et AM s’intéressaient au sevrage tabagique [[47]], fibromyalgie [[48]], leucopénie induite par la chimiothérapie [[49]], l’état dépressif [[50]], la schizophrénie [[51]] et l’obésité [[52]]. Aucune de ces méta-analyses n’étaient concluantes sur une éventuelle supériorité de l’EA par rapport à l’acupuncture manuelle, mais il est bien difficile d’interpréter ces données du fait d’une faible puissance de ces méta-analyses et du fait que les comparaisons étaient indirectes, à l’exception de la méta-analyse sur la gonalgie montrant une efficacité supérieure de l’EA. En attendant de nouvelles comparaisons et en se fiant aux études expérimentales, il apparaît néanmoins que l’EA a un effet supérieur à l’acupuncture manuelle seule.
Effets immédiats de l’acupuncture : mythe ou réalité ?
L’intérêt aussi de cette étude clinique est d’objectiver que l’acupuncture a un effet immédiat, aussi rapide en action que le fait de prendre de l’acide acétylsalicylique pour faire baisser la fièvre ou la douleur.
D’ailleurs, cet effet immédiat de l’acupuncture et techniques associées a pu également être démontré dans diverses pathologies et symptômes : hémiplégie à la suite d’un accident ischémique cérébral ou effets neuromusculaires après AVC [53-55], effets antalgiques dans la gonalgie liée à la gonarthrose [[56],[57]], actions sur la mécanosensitivité du nerf médian [[58]], dans les douleurs oromyofaciales, craniomandibulaires, cervicales [[59],[60]], sur la rigidité artérielle avec baisse de la pression artérielle diastolique [[61]] ou dans la diminution de la résistance vasculaire de l’artère rénale par moxibustion chez les patients atteints d’insuffisance rénale chronique [[62]], dans la diminution de la force segmentaire musculaire visualisée par électromyographie [[63]], dans la paralysie cérébrale spastique avec diminution de la spasticité du tonus du muscle gastrocnemius chez les enfants [[64]], dans les menstruations et les dysménorrhées primaires [[65]], dans le bien-être psychologique, les réponses immunologiques et endocriniennes physiologiques chez le sportif en compétition [[66]], dans l’amélioration de la dyspnée et de la FEV1 de la broncho-pneumopathie chronique obstructive [[67]], dans l’asthme bronchique et les rhinites allergiques chez les enfants et les adolescents [[68],[69]], dans l’analgésie [[70]], dans la dyspepsie fonctionnelle et la variabilité de la fréquence cardiaque [[71]], etc.
Bien sûr ces études ne sont pas toutes des études comparatives randomisées de grande puissance, mais il s’avère donc que l’acupuncture et techniques associées peuvent avoir un effet immédiat dans de nombreuses pathologies.
Conclusion
Comme nous l’avons vu, chez le patient diabétique sous insuline, la variabilité glycémique est directement liée à la perte de l’homéostasie. On peut la mesurer par des outils spécifiques de calcul qui vont permettre d’évaluer les variations intra-journalières de la glycémie à court terme (MAGE), les variations inter-journalières (MODD), etc.. Ainsi, ces deux indices (MAGE et MODD) apprécient chacun une composante spécifique de la variabilité glycémique du patient diabétique insulino-traité. Certains auteurs considèrent que le MAGE est le témoin d’une variabilité endogène (défaut de sécrétion de l’insuline) et le MODD le témoin d’une composante exogène (type de traitement utilisé) de la variabilité glycémique. Le MAGE, coefficient de variation du glucose, défini par la déviation standard (DS) ajustée sur la moyenne glycémique, est certainement le meilleur index pour évaluer la variabilité glycémique. La valeur de 36 % permet de séparer les diabètes stables et labiles [3,[72]].
Par ailleurs, chez le diabétique, la valeur moyenne du MODD est significativement plus élevée chez les patients présentant des changements de mode de vie que chez ceux qui n’en ont pas [[73]].
Dans notre observation, il est difficile de les calculer du fait du nombre insuffisant de données recueillies au domicile (tableau I) [3,[74]]. Néanmoins en raison du mode de vie sédentaire et de son invalidité au travail du patient, on peut supposer que la variabilité glycémique est faible et que son diabète de type 2 est stable. Cela témoigne que l’acupuncture et EA associées au traitement diététique, médicamenteux permet une faible perte de l’homéostasie glycémique, ce qui réduira la survenue d’événements cardio-vasculaires graves chez ce patient diabétique de stade 2 insulino-requérant. L’effet est immédiat et se prolonge dans le temps.
En conclusion, l’acupuncture et l’électroacupuncture, se doivent de faire partie de la panoplie du traitement de tout diabète de type 2.
Points forts
L’EA a un effet hypoglycémiant immédiat dans le diabète de type 2. Protocole de points : 36E zusanli, 12VC zhongwan et 4VC guanyuan à la fréquence 15Hz (durée d’impulsion 200µs) L’EA a un effet hypoglycémiant à long terme dans le diabète de type 2. Protocole de points : 3/80 Hz ; 300µs sur 6MC (neiguan), 6Rt (sanyinjiao) Autres points à utiliser : puncture manuelle avec recherche du deqi sur les points 23V (shenshu), 3R (taixi), 4Rt (gongsun), 7C (shenmen), 20VG (baihui), yishu (EX-B3)
Merisiers en fleurs – Parc Naturel Régional Scarpe Escaut – France
Annexes
Annexe 1. Le choix des points
Les points ont été choisis selon les Textes.
Xiaoke (de xiao amaigrissement et ke soif), diabète de type 2 est lié à une Chaleur des Trois Réchauffeurs (sanjiao) et est dû à un Vide de yin engendrant un feu interne. Dans ce cas clinique, cela a abouti ainsi à un Vide de yin et de yang des Reins. Il est alors intéressant d’utiliser les points choisis ci-dessous.
Neiguan (6MC)
Il s’agit du point clé d’ouverture du yinweimai, Merveilleux Vaisseau qui contrôle qualitativement le yin. Par ailleurs, il est le point luo du shoujueyin permet de faire croître le yin et de stabiliser le shen. Il a pour fonction d’ouvrir la poitrine, calmer le shen, faire circuler le qi et le Sang (xue), le qi du Foie, harmoniser l’Estomac.
Sanyinjiao(6Rt)
Il harmonise et tonifie le qi de la Rate et de l’Estomac ; c’est le point luo de groupe des méridiens yin des membres inférieurs (Rate, Foie, Reins) et de ce fait contribue à remonter le yin du bas de la Rate, des Reins et du Foie. Il élimine l’Humidité, calme le shen, nourrit le Sang et le yin. Associé à guanyuan (4VC), il peut renforcer le xian tian (Inné ou Ciel antérieur), le hou tian (Acquis ou Ciel postérieur), le yuan yang (yang originel) et le yuan yin (yin originel).
Guanyuan (4VC)
Il est le point mu d’Intestin Grêle, point de croisement du Vaisseau Conception avec Rate, Foie et Reins et chongmai. Tonifie le yin des Reins, tonifie le qi Originel (yuan qi). De plus en tonification et réchauffement, 4RM renforce le yang des Reins. Il calme également le shen.
Zusanli (36E)
C’est unpoint Mer (he) du méridien d’Estomac, point Terre, utilisé pour faire descendre le qi et clarifier la Chaleur ; point Étoile Céleste [note 13] de Ma Dan Yang, point de la Mer de l’Eau et des Aliments ; va harmoniser Rate et Estomac, le xue et le qi.
Zhongwan (12VC)
Point mu de l’Estomac, point de réunion (hui) des Entrailles (fu), point de croisement avec les méridiens de l’Intestin grêle, du Triple Réchauffeur et de l’Estomac, il tonifie Estomac et Rate, harmonise le Réchauffeur moyen, régule la circulation du qi, élimine les Mucosités-Glaires (tanyin) et l’Humidité.
Shenshu (23V)
Point beishu (point shu du dos) du Rein, le renforce, tonifie son qi et son yang ; a des effets bénéfiques sur l’Essence (jing), nourrit le yin du Rein.
Taixi (3R)
Point antique (shu), yuan (source) et shu (rivière), point Terre, il tonifie les Reins que ce soit le yin ou le yang des Reins, stabilise le qi du Poumon, élimine la Chaleur-Vide.
Gongsun (4Rt)
Renforce et régule le qi de la Rate car il est son point luo et élimine l’Humidité ; il est aussi le point clé d’ouverture du chongmai.
Shenmen (7C)
Régule et renforce le Cœur (Sang et yin), calme l’Esprit (shen) ; élimine la Chaleur du méridien du Cœur ; Point source (yuan), point rivière (shu), point Terre ; calme l’Esprit (shen).
Baihui (20VG)
Il est le point de croisement du Vaisseau Gouverneur avec les méridiens de Vessie, Vésicule Biliaire, Triple Réchauffeur et du Foie ; point de la Mer des Moëlles. Il a pour action de faire tomber le Vent interne, soumet le yang, a des effets bénéfiques sur le Cerveau et les organes des sens, calme l’Esprit (shen).
Yishu (EX-B3)
C’est un point hors méridien situé à 1,5 cun en dessous épineuse de T8. Cela correspond à la zone métamérique du pancréas.
Annexe 2. Expérimentations animales sur les effets immédiats de l’acupuncture et électroacupuncture
En 1999, Chang et coll. stimulaient pendant 30 mn par EA (fréquence 15Hz, durée d’impulsion 200µs) les points zhongwan (12VC) et gwanyuan (4VC) sur un modèle de rats diabétiques et observaient versus groupe placebo, une baisse de la glycémie statistiquement significative (p<0,01), avec retour de la glycémie à la normale au bout de 30mn après la séance. Les auteurs observaient également une augmentation de la sécrétion plasmatique de β endorphines (p<0,05) et d’insuline (p<0,01) et suggéraient que la réduction de la glycémie était en relation avec ces stimulations. Pas d’effet de l’EA sur le glucagon [6].
Ishizaki et coll. ont démontré sur un modèle de rats diabétiques GK que l’EA (15Hz, 200µs sur 12VC et 4VC) a engendré une hypoglycémie statistiquement significative (p<0,007) mesurée à la 30e mn par augmentation marquée des taux plasmatiques d’insuline (p<0,001) versus groupe témoin. L’EA augmente donc la sensibilité à l’insuline [9].
Après EA (15 Hz ; 200µs) sur zusanli (36E), l’activité hypoglycémique a été confirmée également versus groupes témoins de manière statistiquement significative (p<0,01) à la fois sur les rats Wistar normaux et les rats diabétiques STZ, la glycémie ayant été mesurée avant EA (0 min) et après EA (à 30 et 60 min). Les auteurs observaient aussi une sensibilité accrue à l’insuline [10].
Un effet hypoglycémique de l’EA (2Hz ; 200µs) a été aussi observé par la stimulation du zusanli (36E) chez les rats normaux versus point placebo (p<0,001) associé à une augmentation statistiquement significative 30 mn après EA des niveaux plasmatiques d’insuline (p<0,01) et des β-endorphines (p<0,05). Il n’y avait aucune réponse hypoglycémique de l’EA à 2 Hz EA chez les rats diabétiques STZ. La sérotonine pourrait jouer un rôle dans ce phénomène. En effet, l’injection intraveineuse de sérotonine (5-HT) a diminué la glycémie de manière significative chez les rats normaux. Par conséquent, les auteurs suggéraient que l’EA à 2Hz sur zusanli induirait une réaction hypoglycémique médiée par la sérotonine et les β-endorphines via la sécrétion d’insuline [7].
Sur un modèle de rats en hyperglycémie par ablation de la surrénale (adrénalectomisé) car l’adrénaline inhibe l’insuline, il a été démontré que l’EA à 15hz (200µs) sur zhongwan (12VC) et gwanyuan (4VC) engendre une hypoglycémie significative versus groupe témoin (p<0,01), action inhibée par la naloxone, mesurée après la séance. Ce qui objective la participation des β-endorphines de la glande surrénale dans l’action hypoglycémique de l’EA [8].
On a pu également montrer que l’EA à 15Hz (200µs) sur zusanli pendant 60 mn chez des rats en résistance à l’insuline par injection de prednisolone (SIIR), permet d’améliorer cette résistance à l’insuline en diminuant les acides gras plasmatiques et régulant à la hausse l’expression des protéines de signalisation de l’insuline (IRS1[ note 14] et GLUT4 [note 15]), ce qui améliore l’activité de l’insuline. La baisse de la glycémie et des acides gras libres a été statistiquement observée à 30, 60mn (p<0,05). Par ailleurs, il existe une diminution de l’HOMA-IR (Homeostasis Model Accessment of insuline resistance) dans le groupe EA versus groupe non-EA (p<0,05) [12].
En 2016, on observe encore des résultats sensiblement identiques aussi sur un modèle de rats SIIR induite par un autre stéroïde, le dexaméthasone. L’EA à 15Hz (200µs) sur zusanli pendant 60 minutes engendre également une glycémie et des concentrations en acides gras libres significativement inférieurs à 30/60 minutes versus groupe non traité (p<0,01). De même, l’expression de l’IRS-1 et du GLUT4 a été considérablement augmentée par l’EA (p<0,01). Les auteurs concluent que l’EA augmentent la sensibilité à l’insuline [11].
On obtient à nouveau les mêmes résultats sur l’abaissement du taux de glucose plasmatique à 30, 60 et 90mn après une séance d’EA de 60mn sur zusanli (36E) (15 Hz 200µs) mais cette fois-ci en traitement associé à l’acarbose (ACA inhibiteurs des alpha-glucosidases). Le traitement combiné de l’ACA + EA a eu un effet de réduction du glucose plus important que celui de l’ACA seul chez les rats Wistar normaux (−45% ±3 % contre −19% ± 3%, p< 0,001) et les rats modèles SIIR (−43% ± 2% contre −16% ± 6%, p < 0,001). Ont également été observées dans le groupe ACA + EA des rats SIIR versus groupe témoin, une réduction significative des niveaux des acides gras plasmatiques, une amélioration de l’évaluation du modèle homéostatique de l’indice IR (HOMA-IR) (−48,9% ± 4,0%, p < 0,001) et l’indice de sensibilité à l’insuline (102 % ± 16,9 %, p < 0,001), des augmentations significatives de l’IRS1, du GLUT4 et du récepteur-γ proliférateur activé par le peroxisome (PPAR‐γ) dans le muscle squelettique. Bref, l’EA combinée à l’ACA produit donc un effet plus fort de réduction de glucose que l’ACA seul [14].
Sur des modèles de rats diabétiques STZ, l’EA (15Hz ; 200µs ; 30 mn) aux zusanli (36E) et zhongwan (12VC) diminue l’hyperglycémie analysée 30mn après la séance (p<0,01) versus groupe témoin. L’EA stimule de manière statistiquement significative (p<0,01) l’expression des protéines de signalisation de l’insuline (IRS1, Akt2 [note 16]) qui sont activées par les nerfs cholinergiques du système parasympathique médié par l’acétylcholine et la glande surrénale [15]. Les auteurs confirment ces données en utilisant l’atropine, l’hémicholinium-3 (HC-3) et l’éserine qui sont antagonistes des nerfs cholinergiques et qui vont de ce fait bloquer l’action hypoglycémique de l’EA 15Hz (200µs). Par ailleurs, aucune différence significative dans les valeurs d’insuline ou de concentrations en acides gras libres n’a été notée entre les deux groupes ; toutefois, on observe aussi une diminution de l’HOMA-IR dans le groupe EA (p<0,05) [16]. Même constat dans une autre étude de Lin et coll. On retrouve également des taux significativement plus bas de la glycémie lors de l’EA (15Hz ; 200µs : 36E) versus groupe témoin à 15, 30, 60 et 90 min (p<0,005). Les auteurs objectivent également l’action de l’EA sur les nerfs cholinergiques, sur le neurotransmetteur monoxyde d’azote ou oxyde nitrique (NO) dont la biosynthèse est sous la dépendance, entre autres de l’isoforme neuronale de l’oxyde nitrique synthase (nNOS1)[note 17]. Ils concluent que l’action de l’EA a un effet combiné sur les nerfs cholinergiques et sur l’activité nNOS. Celle-ci est augmentée significativement (p<0,05), via des concentrations plus faibles en acides gras libres (p<0,05 à 90mn) plasmatiques avec amélioration de l’activité de l’insuline par une régulation positive de l’IRS1 dans le muscle squelettique du rat [13].
Annexe 3. Expérimentations animales sur les effets à long terme de l’acupuncture et électroacupuncture
L’axe hypothalamo-hypophyso-surrénalien libère le cortisol qui est augmenté lors d’une période de stress, engendrant la libération d’acides gras et de triglycérides. Cela va diminuer la sensibilité à l’insuline avec le risque de diabète de type 2. Gao et coll. ont ainsi objectivé que l’EA 15Hz (tous les jours 20 min ; 5 traitements par semaine pendant un mois) sur un modèle de rats diabétiques appliquée au point yishu (EX-B3) réduit significativement l’hyperglycémie (p<0,01) tout comme pishu (V20) et shenshu (V23). L’EA atténue l’hyperactivité de l’axe hypothalamo-hypophyso-surrénalien en réduisant la corticotropin-releasing hormone (CRH) et le cortisol, en améliorant la résistance à l’insuline et l’indice de sensibilité à l’insuline et en régulant le métabolisme des lipides sanguins de manière significative (triglycérides, cholestérol HDL et LDL) [21].
L’étude de Tominaga et coll [22] montre que l’application de quatorze séances réparties sur quatre semaines d’EA (2/15Hz ; 200µs) au 36E zusanli engendre une augmentation significative dans le muscle de la protéine kinase activée par l’AMP (AMPK) [note 18] juste après la séance finale versus groupe témoin (p<0,05), donc améliorant la résistance à l’insuline induite par l’alimentation.
Ainsi sur un modèle de rats mâles Sprague-Dawley avec insulinorésistance induite par un régime hyperlipidique (n=24), Lan et coll. ont étudié l’effet de l’EA (2/50Hz ; 500µs ; séance de 20mn tous les jours pendant 4 semaines) sur les points shenshu (V23), neiguan (6MC), zusanli (36E) et sanyinjiao (Rt6). Ils ont observé une augmentation significative des marqueurs de l’expression des IRS1, Pi3K [note 19], Akt2 et de l’eNOS dans l’endothélium vasculaire avec production de monoxyde d’azote (NO) versus groupe témoin ou groupe insulinorésistant non traité. De même l’indice HOMA était significativement abaissé (p<0,05) [25]. Ceci permet d’atténuer le dysfonctionnement endothélial et pourrait donc jouer un rôle potentiel dans la prévention ou le traitement des complications diabétiques communes telles que neuropathies, néphropathies et maladies cardiovasculaires par la voie de signalisation Pi3K/Akt, tout en améliorant l’impact négatif de l’alimentation [17].
Une autre étude expérimentale similaire objective également chez un modèle de rat en insulinorésistance (n=32) que l’EA (2/50Hz ; 500µs ; séance de 20mn sept jours par semaine pendant deux semaines consécutives) sur les points 40E (fenglong) et 6Rt (sanyinjiao) améliore la sensibilité à l’insuline (indice HOMA-IR diminué p<0,01) par régulation positive de l’expression des composants de la signalisation AMPK et de l’acétyl-CoA carboxylase (ACC) [note 20] dans le muscle squelettique versus groupe témoin. Diminution aussi des triglycérides et du cholestérol (p<0,05) [23].
Que l’on retrouve également dans cette étude expérimentale sur quarante rats qui a observé l’effet de l’électroacupuncture (2 Hz/100 Hz) sur les points fenglong (40E) et sanyinjiao (6Rt) dans les troubles du métabolisme des lipides, la résistance à l’insuline (RI) et l’expression des protéines SREBP-1 (sterol regulatory element blinding protein-1) [note 21] au niveau du tissu hépatique de rats atteints d’hyperlipidémie avec RI. Ainsi après EA une fois par jour pendant 14 jours, il est observé une régulation de la glycémie à 60 et 120 mn après dernière intervention, une augmentation significative du cholestérol HDL sérique (P<0,01), une diminution des triglycérides sériques, du cholestérol LDL, des acides gras libres (p<0,05) et des niveaux d’expression des protéines hépatiques SREBP-1c par rapport au groupe témoin (P<0,01). Aucune différence significative n’a été constatée entre les groupes EA et médicaments dans tous les indices mentionnés ci-dessus (P>0,05). En conclusion, l’EA est capable d’améliorer le trouble du métabolisme des lipides des rats en insulinorésistance, ce qui peut être associé à ses effets de réduction de l’expression des protéines SREBP-1 c et de la diminution de la synthèse des acides gras [26]. Les mêmes auteurs, dans une étude similaire avec la même méthodologie d’EA objectiveront une amélioration dans le tissu hépatique du métabolisme lipidique des rats en insulinorésistance en rapport avec une diminution de l’activité des enzymes liées à la synthèse des acides gras, régulation de l’adiponectine et de la leptine [note 22] induite par la régulation de la signalisation AMPK/ p38 MAPK/PPARγ (kinase protéique activée par le mitogène p38/récepteur gamma activé par proliférateur peroxisome (PPARγ) [24].
L’étude de Peplow concernant des rats Zucker, obèses et diabétiques âgés appliquant le protocole précédemment établi [19] (EA 10Hz ; 200µs ; 30mn) a objectivé une diminution de la leptine et une augmentation du rapport adiponectine/leptine dans le tissu adipeux et le sérum. De tels changements peuvent améliorer la résistance à l’insuline chez les rats diabétiques obèses. Cet effet hypoglycémique de l’EA répétée sur zhongwan (12VC) et guanyuan (4VC) est plus efficace que l’EA sur zusanli (36E). La diminution de la leptine peut ainsi améliorer la sensibilité à l’insuline et augmenter l’absorption de glucose stimulée par l’insuline dans les adipocytes [28].
L’EA (2/15Hz ; 500µs) ; 30mn sur baihui (20VG) a également un effet protecteur sur les lésions ischémiques cérébrales chez les souris diabétiques STZ, lorsqu’elle est utilisée deux heures avant induction d’une ischémie cérébrale focale. Les auteurs observent une réduction de la taille de l’infarctus, une diminution du malondialdéhyde (MDA) [note 23] et des espèces réactives d’oxygène (ROS) [note 24]. Cela est médié au moins en partie par l’action inhibitrice de l’EA de l’activité oxydase de la NADPH. Les résultats suggèrent que l’EA peut avoir un rôle potentiel dans la prophylaxie des complications cérébro-vasculaires du diabète [29].
De nombreuses études ont montré aussi que l’épigénétique est étroitement associée à l’obésité mais également aux troubles métaboliques [32,33]. L’acupuncture ou/et l’EA peuvent avoir aussi un rôle épigénétique dans le diabète de type 2 comme cela a été démontré dans le stress [[75]].
Ainsi, par le biais de la déméthylation et l’inhibition du promoteur hypothalamique Tsc1(Tuberous sclerosis complex 1), l’EA (2/15hz ; durée impulsion 1000µs ; 30mn/jour pendant trente jours) sur tianshu 25E, zhongwan 12VC, sanyinjiao 6Rt et zusanli 36E va ainsi inhiber l’activité de la voie de signalisation mTORC1 [note 25], comme cela a été démontré expérimentalement chez des rats en obésité par alimentation en haute teneur de matières grasses. L’EA contrôle ainsi les facteurs de régulation de l’appétit et atténue l’obésité chez ces animaux [31].
Une nouvelle approche des mécanismes physiopathologiques de l’action de l’EA sur le diabète fait intervenir l’action sur les ARN circulaires [note26] [33] par une autre étude épigénétique qui va conforter les résultats de l’étude de Leng et coll. [31].
Ainsi il est observé après EA (2Hz pendant 15 mn ; durée d’impulsion 500µs) sur zusanli (36E), pishu (20V) une fois tous les deux jours, trois fois par semaine, pendant 4 semaines (12 fois au total) une réduction de la glycémie à jeun, une préservation de la structure de l’îlot et une réduction du taux apoptotique de cellules β de l’îlot de Langerhans chez les souris modèles de diabète de type 2. Par ailleurs, cent-soixante-cinq circARN exprimés différemment ont été retrouvés [note 27]. Ces résultats suggèrent que l’action de l’EA est multifactorielle. La régulation de la glycémie dans le diabète de type 2, peut aussi se faire éventuellement via la régulation de la signalisation de l’hormone thyroïdienne et du phosphatidyl-inositol [34].
Notes
[1]. Si l’IMC est : Entre 25,0 et 29,9 kg/m², il existe un surpoids ; Entre 30,0 et 34,9 kg/m², il s’agit d’obésité modérée ; Entre 35,0 et 39,9 kg/m², il s’agit d’une obésité sévère ; Plus de 40 kg/m², on parle d’obésité massive.
[2]. Un test de glycémie aléatoire peut être effectué à tout moment de la journée. Les résultats peuvent varier en fonction du dernier repas consommé. On considère cependant qu’une personne en bonne santé avec une tolérance au glucose normale n’aura pas de changements majeurs dans les niveaux de glucose dans le sang. Un taux de glucose aléatoire normal varie de 0,70g/L (3,9mmol/L) et ne dépasse pas 1,40g/L (7,8mmol/L).
[3]. La streptozotocine, à forte dose, détruit les cellules B ; à faible dose et répétée, induit une destruction des cellules B de Langerhans par un mécanisme immunitaire dépendant des cellules T.
[4]. Dans son milieu naturel, cet animal se nourrit de plantes salées pauvres en calories, alors que soumis à un régime standard de laboratoire, 40% des animaux deviennent obèses et développent un diabète non insulino-dépendant à partir du 3e mois. Les 60% restants ne présentent pas de diabète mais restent obèses avec des taux élevés d’insuline plasmatique. Le rat des sables répond à l’augmentation alimentaire provoquant une surcharge calorique par un accroissement du poids corporel dû à une augmentation de la taille des adipocytes ou cellules graisseuses, une hyperinsulinémie et une intolérance au glucose à différents degrés.
[5]. L’épigénome comprend la méthylation de l’ADN, les modifications de l’histone et les ARN non codants, qui peuvent réguler la différenciation cellulaire, l’expression des gènes spécifiques aux cellules, l’empreinte parentale, l’inactivation du chromosome X, ainsi que la stabilité et la structure génomiques [30].
[6]. Dans les études expérimentales, la fréquence la plus souvent appliquée est 2/50 Hz avec une durée d’impulsion de 500µs, ce qui correspond à la fréquence 3/80 Hz à une durée d’impulsion 300µs.
[7]. Cela correspond à une fréquence de 1/40 Hz pour une durée d’impulsion 200µs
[8]. Zhongping est localisé à mi-distance entre le 38E et l’interligne fémoro-tibial. Le point est décrit comme un point curieux sous le nom de zuzhongping (zhongping du pied). Voir Goret O. Tiaokou dans l’épaule douloureuse : shang bing xia qu. Acupuncture & Moxibustion. 2005;4(3):218-219.
[9]. Une valeur I² <25% indique une hétérogénéité faible, des valeurs comprises entre 25 et 50% une hétérogénéité modérée et une valeur 50% une hétérogénéité importante.
[10]. L’insulinorésistance se définit par une réduction de la réponse biologique à l’action de l’insuline. De ce fait, on observe des anomalies du métabolisme des glucides, pouvant conduire à l’apparition d’un diabète de type 2 ; des anomalies des lipides (hypertriglycéridémie, hypercholestérolémie), et est souvent associée à une surcharge pondérale et à une hypertension artérielle.
[11]. L’HOMA-IR est un index qui permet de calculer la résistance à l’insuline (insulino-résistance). HOMA-IR = Insuline x Glucose / 22,5. Les patients ayant un index de HOMA > 3 sont insulino-résistants
[12]. Le syndrome métabolique ne se définit pas comme étant une maladie, mais plutôt se traduit par la présence de plusieurs troubles physiologiques et biochimiques. Reconnu depuis les années 1920, le syndrome métabolique, aussi appelé « syndrome X », désigne la coexistence de plusieurs désordres métaboliques qui augmentent considérablement le risque de diabète de type 2 et de maladies cardiaques. Le diagnostic est fait lorsque le sujet présente au moins trois des symptômes suivants : hypertension artérielle, glycémie élevée mais sans réel diabète de type 2, surpoids ou embonpoint abdominal, faible taux de HDL cholestérol et taux élevé de triglycérides. Ces signes peuvent entraîner des complications (passage en diabète de type 2, cardiopathie, d’accident vasculaire cérébral (AVC). Bref, cela est engendré par une insulino-résistance. La prévalence du syndrome métabolique en France est de 22,5% chez les hommes et 18,5% chez les femmes selon l’étude de Gamila et al. (Gamila S, Dallongeville J. Épidémiologie du syndrome métabolique en France ; 2003.)
[13]. Les points Etoiles célestes ont été tout d’abord onze décrits par Ma Dan Yang (1123-1183) dans sa « chanson des onze points Etoiles célestes ». Le douzième point taichong (F3) a été rajouté sous la dynastie Ming (1368-1644) par Xu Feng Zhuan dans son « Classique du Dragon de Jade ». Aujourd’hui on les retrouve dans le classique Zhenjiu Dacheng (针灸大成) daté de 1601. Les points sont : E36 zusanli, E44 neiting, GI11 quchi, GI4 hegu, V40 weizhong, V57 chengshan, F3 taichong, V60 kunlun, VB30 huantiao, VB34 yanglingquan, C5 tongli, P7 lieque.
[14]. Le substrat récepteur de l’insuline 1 (IRS1) est une protéine adaptatrice de signalisation. La transmission du signal insulinique dans la cellule met en jeu des modules protéiques de reconnaissance présents sur les protéines substrats et capables de les positionner à proximité du récepteur activé. L’IRS1 joue un rôle clé dans la transmission de signaux de l’insuline et l’insuline-like growth factor-1 (IGF-1), mais aussi des récepteurs aux voies intracellulaires Pi3K/Akt et les voies Erk MAP Kinases
[15]. Le transporteur de glucose 4 (GLUT4) est une protéine de la famille des transporteurs GLUT de classe I (avec GLUT1, GLUT2, et GLUT3), dont le rôle essentiel est le transport du glucose du plasma au muscle et au tissu adipeux.
[16]. L’Akt2 est une enzyme appartenant à la famille des protéines kinases, impliquée dans la signalisation cellulaire de l’insuline, nécessaire au transport du glucose.
[17]. Le nNOS est exprimé dans les îlots pancréatiques de rat et aide à contrôler la sécrétion d’insuline.
[18]. La protéine kinase activée par l’AMP (AMPK) est une protéine régulatrice clé de l’homéostasie énergétique et joue entre autres dans la régulation de la synthèse d’insuline. L’AMPK contenu dans le muscle squelettique va médier également le transporteur insulino-dépendant du glucose GLUT4.
[19]. L’une des principales voies de la signalisation insulinique est celle de la phosphatidyl-inositol 3 kinase (Pi3K). Dans la voie de signalisation PI3K/Akt, l’activité du Pi3K des rats résistants à l’insuline est généralement inhibée. De ce fait, les activités métaboliques comme la synthèse du glycogène sont améliorées via la voie de signalisation Pi3K/Akt. (Capeau J. Voies de signalisation de l’insuline : mécanismes affectés dans l’insulino-résistance. M/S : médecine sciences. 2003;19(8–9) :834-839).
[20]. L’acétyl-CoA carboxylase est une enzyme largement répandue dans de nombreux tissus mais principalement dans le foie, le tissu adipeux et la glande mammaire qui synthétisent des acides gras. Elle intervient dans la première étape de la synthèse des acides gras. L’insuline produit l’induction de la transcription du gène de l’acétyl-CoA carboxylase.
[21]. Le SREBP-1c est un médiateur des effets transcriptionnels de l’insuline dans le foie, acteur majeur de la régulation du métabolisme lipidique puisqu’ils contrôlent l’expression des gènes du métabolisme des acides gras, des triglycérides et du cholestérol. L’activation de l’isoforme SREBP-1c se fait par l’insuline dans le foie. L’insuline après liaison à son récepteur active la transcription du gène SREBP-1c par une voie impliquant la phosphatidyl inositol 3 phosphate kinase (PI3 kinase) et la protéine kinase B (PKB). (Foufelle F, Hegarty B, Bobard A, Hainault I, Bossard P, Ferré P. Un nouveau rôle de l’insuline dans la régulation du métabolisme glucido-lipidique hépatique. Médecine/scienses.2005;21:569-92.).
[22]. L’adiponectine est produite par le tissu adipeux et impliquée, entre autres, dans la régulation du métabolisme des lipides et du glucose. Un taux d’adiponectine élevé diminuerait le risque de diabète de type II. Elle exerce son rôle antidiabétique au niveau du foie et du muscle squelettique en augmentant la sensibilité à l’insuline de ces organes. Au niveau hépatique, elle permet de diminuer la production de glucose et de réduire le contenu en triglycérides. Dans le muscle squelettique, l’adiponectine permet une augmentation de l’entrée de glucose et une augmentation de l’oxydation des acides gras. La leptine est produite et sécrétée dans la circulation sanguine par les adipocytes. Une sécrétion importante de leptine induit une lipolyse, inhibe la lipogenèse et augmente la sensibilité à l’insuline. La leptine agirait comme un signal de satiété. (Vatier C ; Capeau J, Vigouroux C. La leptine, nouvelle perspective thérapeutique dans le traitement du diabète ? Médecine/Sciences. 2010 ;10(26):504-807)
[23]. Le stress oxydant peut être évalué par la mesure du produit terminal de la péroxidation lipidique, le malondialdéhyde (MDA). C’est un marqueur de stress oxydant sensible et spécifique.
[24]. La NADPH oxydase (NOX) est un complexe enzymatique membranaire, enzyme clé de l’explosion oxydative, où elle catalyse la réaction d’oxydation du NADPH par l’oxygène O2, ce qui produit du NADP+, de l’H+ et de l’O2–. Ces deux derniers réagissent ensuite sous l’action de la superoxyde dismutase (SOD) pour former du peroxyde d’hydrogène H2O2.
[25]. Parmi les nombreuses voies de signalisation de la cellule cancéreuse, la voie PI3K-Akt-mTOR joue un rôle majeur dans la croissance, la prolifération et la survie cellulaire. Il s’agit d’une voie complexe dont l’activation peut se faire soit au niveau extracellulaire via des récepteurs à activité tyrosine kinase, soit au niveau intracellulaire par mutation ou surexpression de protéines impliquées dans cette voie. mTOR (mechanistic target of rapamycin) dans l’hypothalamus, cible de la rapamycine chez les mammifères, est une enzyme qui régule donc la prolifération, la croissance, la mobilité cellulaire, etc. De ce fait, elle est associée au niveau pathologique à la genèse des tumeurs. Cependant, mTOR joue aussi un rôle dans l’obésité, la faim et l’homéostasie énergétique de l’organisme. On objectivera ainsi sur des modèles de rats obèses une augmentation de la méthylation du promoteur du gène Tsc1 et celle de l’expression du mTORC1 dans l’hypothalamus. La voie de signalisation mTORC1 est activée par les nutriments, les hormones comme l’insuline et les facteurs de croissance apparentés à l’insuline 1 (IGF-1) par le biais de la voie PI3K/Akt ou de la voie des MAPK, et aussi nécessite l’inhibition de Tsc1/Tsc2 au niveau hypothalamique. L’AMPK inhibe mTORC1 dans des conditions de déficit en énergie.
[26]. Les ARN circulaires (circARN) sont actuellement classés comme des ARN non codants (ARNnc), biomarqueurs potentiels de la maladie. Des exosomes, nanovésicules extracellulaires (30–150 nm) d’origine endocytique transportent de nombreuses molécules biologiques telles que ces fragments d’ADN, des circARN et des micro(mi)ARN qui favorisent la communication intercellulaire et régulent de nombreux processus pathophysiologiques. Ainsi une surexpression du miR-7 (miARN-7) dans les cellules β transgéniques de souris entraîne le diabète. De même, il a également été démontré que la surexpression du ciRS-7 inhibe la fonction miR-7 dans les cellules β de l’îlot de Langerhans, ce qui améliore à son tour la sécrétion d’insuline. Notons qu’il existe plusieurs couches de régulation épigénétique : méthylation directe des résidus de cytosine ou d’adénine, modifications covalentes des protéines histones, structure chromatine de plus haut ordre et ARN non codant, comme ici. Chacun d’entre eux a été impliqué dans des processus cellulaires intervenant dans le diabète. De fait, il existe de nombreuses associations entre l’épigénétique et le diabète, l’obésité, et autres désordres métaboliques.
[27]. Shou et coll. montrent dans leur étude que l’EA peut agir à plusieurs niveaux : réguler à la hausse les voies de signalisation de l’hormone thyroïdienne, celles de la régulation des erreurs de transcription du cancer, de la régulation des lipides des adipocytes, etc. ; et peut réguler à la baisse la voie de transduction du signal de la phagocytose des macrophages, la voie du système de signalisation du phosphatidyl-inositol, la voie du métabolisme de la choline cancéreuse, la voie de signalisation mTOR, la voie du métabolisme du phosphoinositol (IP) et la voie du protéoglycane cancéreux, etc. Ainsi, les analyses ont révélé entre autres que la signalisation de l’hormone thyroïdienne était activement régulée par l’EA. L’analyse de l’interaction circARN/miARN a révélé aussi que le mmu-mir-7092-3p était étroitement associé au circINPP4B, suggérant que la voie de signalisation du phosphatidyl-inositol pouvait être affectée par l’EA.
Références
[1]. Inserm et Burcelin R (unité 1048 Inserm/Université Toulouse-3 Paul Sabatier). Diabète de type 2. Un trouble du métabolisme principalement lié au mode de vie. Février 2019. [consulté le 18/07/2020]. Disponible à l’URL : https://www.inserm.fr/information-en-sante/dossiers-information/diabete-type-2
[2]. Haute Autorité de Santé (HAS). Recommandations de bonne pratique portant sur la stratégie médicamenteuse du contrôle glycémique du diabète de type 2. Janvier 2013. [Consulté le 12/07/2020]. Disponible à l’URL : https://www.has-sante.fr/upload/docs/application/pdf/2013-02/10irp04_reco_diabete_type_2.pdf.
[3]. Guerci B. La variabilité glycémique asymptomatique : comment l’évaluer et quelle est son incidence clinique ? [Asymptomatic glycemic instability: how to measure it and which clinical applications? ]. Diabetes Metab. 2003 Apr;29(2 Pt 1):179-88.
[4]. Hawawini R. Le diabète en acupuncture, en dehors des complications : à propos de deux formules thérapeutiques. Acupuncture & Moxibustion. 2017;16(2) :139-144.
[5]. BiostaTGV. Site de biostatistiques en ligne. [consulté le 18/07/2020]. Disponible à l’URL : https://biostatgv.sentiweb.fr/.
[6]. Chang SL, Lin JG, Chi TC, Liu IM, Cheng JT. An insulin-dependent hypoglycaemia induced by electroacupuncture at the Zhongwan (CV12) acupoint in diabetic rats. Diabetologia. 1999;42(2):250-5.
[7]. Chang SL, Tsai CC, Lin JG, Hsieh CL, Lin RT, Cheng JT. Involvement of serotonin in the hypoglycemic response to 2 Hz electroacupuncture of zusanli acupoint (ST36) in rats. Neurosci Lett. 2005;379(1):69-73.
[8]. Lin JG, Chen WC, Hsieh CL, Tsai CC, Cheng YW, Cheng JT, Chang SL. Multiple sources of endogenous opioid peptide involved in the hypoglycemic response to 15 Hz electroacupuncture at the Zhongwan acupoint in rats. Neurosci Lett. 2004;366(1):39-42.
[9]. Ishizaki N, Okushi N, Yano T, Yamamura Y. Improvement in glucose tolerance as a result of enhanced insulin sensitivity during electroacupuncture in spontaneously diabetic Goto-Kakizaki rats. Metabolism. 2009 Oct;58(10):1372-8.
[10]. Chang SL, Lin KJ, Lin RT, Hung PH, Lin JG, Cheng JT. Enhanced insulin sensitivity using electroacupuncture on bilateral Zusanli acupoints (ST 36) in rats. Life Sci. 2006;79(10):967-71.
[11]. Tzeng CY, Lee YC, Chung JJ, Tsai JC, Chen YI, Hsu TH, Lin JG, Lee KR, Chang SL. 15 Hz electroacupuncture at ST36 improves insulin sensitivity and reduces free fatty acid levels in rats with chronic dexamethasone-induced insulin resistance. Acupunct Med. 2016;34(4):296-301.
[12]. Lin RT, Tzeng CY, Lee YC, Ho WJ, Cheng JT, Lin JG, Chang SL. Acute effect of electroacupuncture at the Zusanli acupoints on decreasing insulin resistance as shown by lowering plasma free fatty acid levels in steroid-background male rats. BMC Complement Altern Med. 2009;9:26.
[13]. Lin RT, Chen CY, Tzeng CY, Lee YC, Cheng YW, Chen YI, Ho WJ, Cheng JT, Lin JG, Chang SL. Electroacupuncture improves glucose tolerance through cholinergic nerve and nitric oxide synthase effects in rats. Neurosci Lett. 2011;494(2):114-118.
[14]. Chung YC, Chen YI, Lin CM, Chang SW, Hsu TH, Ho WJ, Lin JG, Chang SL, Tzeng CY. Electroacupuncture combined with acarbose improves insulin sensitivity via peroxisome proliferator-activated receptor γ activation and produces a stronger glucose-lowering effect than acarbose alone in a rat model of steroid-induced insulin resistance. Acupunct Med. 2020 Oct;38(5):335-342.
[15]. Lee YC, Li TM, Tzeng CY, Chen YI, Ho WJ, Lin JG, Chang SL. Electroacupuncture at the Zusanli (ST-36) Acupoint Induces a Hypoglycemic Effect by Stimulating the Cholinergic Nerve in a Rat Model of Streptozotocine-Induced Insulin-Dependent Diabetes Mellitus. Evid Based Complement Alternat Med. 2011;2011:650263.
[16]. Lee YC, Li TM, Tzeng CY, Cheng YW, Chen YI, Ho WJ, Lin JG, Chang SL. Electroacupuncture-induced cholinergic nerve activation enhances the hypoglycemic effect of exogenous insulin in a rat model of streptozotocin-induced diabetes. Exp Diabetes Res. 2011;2011:947138.
[17]. Feng Y, Fang Y, Wang Y, Hao Y. Acupoint Therapy on Diabetes Mellitus and Its Common Chronic Complications: A Review of Its Mechanisms. Biomed Res Int. 2018 Oct 22;2018:3128378.
[18]. Shapira MY, Appelbaum EY, Hirshberg B, Mizrahi Y, Bar-On H, Ziv E. A sustained, non-insulin related, hypoglycaemic effect of electroacupuncture in diabetic Psammomys obesus. Diabetologia. 2000;43(6):809-13.
[19]. Peplow PV, Han SM. Repeated application of electroacupuncture ameliorates hyperglycemia in obese Zucker diabetic fatty rats. J Acupunct Meridian Stud. 2014 Feb;7(1):1-5..
[20]. Peplow PV, Baxter GD. Electroacupuncture for control of blood glucose in diabetes: literature review. J Acupunct Meridian Stud. 2012 Feb;5(1):1-10.
[21]. Gao S, Li R, Tian HH, Pei ES, Cao BY, Wu Y. [Effects of electroacupuncture at « Yishu » (EX-B 3) on the relative hormones of HPA axis in rats with type-2 diabetes mellitus]. Zhongguo Zhen Jiu. 2014 ;34(11):1099-105.
[22]. Tominaga A, Ishizaki N, Naruse Y, Kitakoji H, Yamamura Y. Repeated application of low-frequency electroacupuncture improves high-fructose diet-induced insulin resistance in rats. Acupunct Med. 2011 Dec;29(4):276-83.
[23]. Li Z, Lan D, Zhang H, Zhang H, Chen X, Sun J. Electroacupuncture Mitigates Skeletal Muscular Lipid Metabolism Disorder Related to High-Fat-Diet Induced Insulin Resistance through the AMPK/ACC Signaling Pathway. Evid Based Complement Alternat Med. 2018 Nov 7;2018:7925842..
[24]. Li ZX, Zhang HH, Lan DC, Zhang HT, Sun J. [Eletroacupuncture improves lipid metabolic disorder by regulating hepatic AMPK/p38 MAPK/RRARγ signaling in rats with high-fat diet-induced insulin resistance]. Zhen Ci Yan Jiu. 2019 Jan 25;44(1):8-12.
[25]. Lan D, Xu N, Sun J, Li Z, Liao R, Zhang H, Liang X, Yi W. Electroacupuncture mitigates endothelial dysfunction via effects on the PI3K/Akt signalling pathway in high fat diet-induced insulin-resistant rats. Acupunct Med. 2018 Jun;36(3):162-169.
[26]. Li ZX, Zhang HH, Lan DC, Zhang HT, Chen XZ, Sun J. [Effect of Eletroacupuncture Intervention on Insulin Resistance, Lipid Metabolic Disorder and Expression of Hepatic SREBP-1 c and Fatty Acid Synthase Proteins in Rats with Hyperlipidemia]. Zhen Ci Yan Jiu. 2018 Jan 25;43(1):8-13.
[27]. Liu X, He J, Qu Y, Liu Z, Hu Y, Guo S, Du J, Jiang P. [Effects of electroacupuncture on insulin resistance and hypothalamic insulin signal molecule in rats with diet-induced obesity]. Zhongguo Zhen Jiu. 2016 Sep 12;36(9):957-961.
[28]. Peplow PV. Repeated electroacupuncture in obese Zucker diabetic fatty rats: adiponectin and leptin in serum and adipose tissue. J Acupunct Meridian Stud. 2015 Apr;8(2):66-70.
[29]. Guo F, Song W, Jiang T, Liu L, Wang F, Zhong H, Yin H, Wang Q, Xiong L. Electroacupuncture pretreatment inhibits NADPH oxidase-mediated oxidative stress in diabetic mice with cerebral ischemia. Brain Res. 2014 Jul 21;1573:84-91.
[30]. Greene J, Baird AM, Brady L, Lim M, Gray SG, McDermott R, Finn SP. Circular RNAs: Biogenesis, Function and Role in Human Diseases. Front Mol Biosci. 2017 Jun 6;4:38.
[31]. Leng J, Xiong F, Yao J, Dai X, Luo Y, Hu M, Zhang L, Li Y. Electroacupuncture Reduces Weight in Diet-Induced Obese Rats via Hypothalamic Tsc1 Promoter Demethylation and Inhibition of the Activity of mTORC1 Signaling Pathway. Evid Based Complement Alternat Med. 2018 Apr 26;2018:3039783.
[32]. van Dijk SJ, Tellam RL, Morrison JL, Muhlhausler BS, Molloy PL. Recent developments on the role of epigenetics in obesity and metabolic disease. Clin Epigenetics. 2015 Jul 11;7:66.
[33]. Greene J, Baird AM, Brady L, Lim M, Gray SG, McDermott R, Finn SP. Circular RNAs: Biogenesis, Function and Role in Human Diseases. Front Mol Biosci. 2017 Jun 6;4:38.
[34]. Shou Y, Hu L, Zhang W, Gao Y, Xu P, Zhang B. Determination of Electroacupuncture Effects on circRNAs in Plasma Exosomes in Diabetic Mice: An RNA-Sequencing Approach. Evid Based Complement Alternat Med. 2019 Sep 24;2019:7543049.
[35]. Belivani M, Lundeberg T, Cummings M, Dimitroula C, Belivani N, Vasilakos D, Hatzitolios A. Immediate effect of three different electroacupuncture protocols on fasting blood glucose in obese patients: a pilot study. Acupunct Med. 2015;33(2):110-4.
[36]. Mohanty S, Mooventhan A, Manjunath NK. Effect of Needling at CV-12 (Zhongwan) on Blood Glucose Levels in Healthy Volunteers: A Pilot Randomized Placebo Controlled Trial. J Acupunct Meridian Stud. 2016;9(6):307-310.
[37]. Kumar R, Mooventhan A, Manjunath NK. Immediate Effect of Needling at CV-12 (Zhongwan) Acupuncture Point on Blood Glucose Level in Patients with Type 2 Diabetes Mellitus: A Pilot Randomized Placebo-Controlled Trial. J Acupunct Meridian Stud. 2017;10(4):240-244.
[38]. Mooventhan A, Ningombam R, Nivethitha L. Effect of bilateral needling at an acupuncture point, ST-36 (Zusanli) on blood glucose levels in type 2 diabetes mellitus patients: A pilot randomized placebo controlled trial [published online ahead of print, 2020 May 14]. J Complement Integr Med. 2020;/j/jcim.ahead-of-print/jcim-2019-0100/jcim-2019-0100.xml. doi:10.1515/jcim-2019-0100.
[39]. Chen C, Liu J, Sun M, Liu W, Han J, Wang H. Acupuncture for type 2 diabetes mellitus: A systematic review and meta-analysis of randomized controlled trials. Complement Ther Clin Pract. 2019 Aug;36:100-112.
[40]. Wu L, Chen X, Liu Y, Lan J, Wu C, Li Z, Lu L, Yi W. Role of acupuncture in the treatment of insulin resistance: A systematic review and meta-analysis. Complement Ther Clin Pract. 2019 Nov;37:11-22.
[41]. Song AQ, Zhang YP, Chen R, Liang FX. Is Acupuncture Effective for Improving Insulin Resistance? A Systematic Review and Meta-analysis. Curr Med Sci. 2018 Dec;38(6):1109-1116.
[42]. Kim TH, Choi TY, Shin BC, Lee MS. Moxibustion for managing type 2 diabetes mellitus: a systematic review. Chin J Integr Med. 2011 Aug;17(8):575-9. doi: 10.1007/s11655-011-0811-2. Epub 2011 Aug 9. PMID: 21826591.
[43]. Li X, Jia HX, Yin DQ, Zhang ZJ. Acupuncture for metabolic syndrome: systematic review and meta-analysis. Acupunct Med. 2020 Oct 9:964528420960485. doi: 10.1177/0964528420960485. Epub ahead of print. PMID: 33032446.
[44]. Langevin HM, Schnyer R, MacPherson H, Davis R, Harris RE, Napadow V, Wayne PM, Milley RJ, Lao L, Stener-Victorin E, Kong JT, Hammerschlag R; Executive Board of the Society for Acupuncture Research. Manual and electrical needle stimulation in acupuncture research: pitfalls and challenges of heterogeneity. J Altern Complement Med. 2015;21(3):113-28
[45]. Mackenzie IZ, Xu J, Cusick C, Midwinter-Morten H, Meacher H, Mollison J, Brock M. Acupuncture for pain relief during induced labour in nulliparae: a randomised controlled study. BJOG. 2011;118(4):440-7.
[46]. Manheimer E, Cheng K, Linde K, Lao L, Yoo J, Wieland S, van der Windt DA, Berman BM, Bouter LM. Acupuncture for peripheral joint osteoarthritis. Cochrane Database Syst Rev. 2010
[47]. White AR, Rampes H, Liu JP, Stead LF, Campbell J. Acupuncture and related interventions for smoking cessation. Cochrane Database Syst Rev 2011;(1):CD000009.
[48]. Langhorst J, Klose P, Musial F, Irnich D, Häuser W. Efficacy of acupuncture in fibromyalgia syndrome–a systematic review with a meta-analysis of controlled clinical trials. Rheumatology (Oxford). 2010 Apr;49(4):778-88.
[49]. Lu W, Hu D, Dean-Clower E, Doherty-Gilman A, Legedza AT, Lee H, Matulonis U, Rosenthal DS. Acupuncture for chemotherapy-induced leukopenia: exploratory meta-analysis of randomized controlled trials. J Soc Integr Oncol. 2007;5(1):1-10.
[50]. Smith CA, Hay PP, Macpherson H. Acupuncture for depression. Cochrane Database Syst Rev. 2010 Jan 20;(1):CD004046. doi: 10.1002/14651858.CD004046.pub3. Update in: Cochrane Database Syst Rev. 2018 Mar 04;3:CD004046.
[51]. Lee MS, Shin BC, Ronan P, Ernst E. Acupuncture for schizophrenia: a systematic review and meta-analysis. Int J Clin Pract. 2009 Nov;63(11):1622-33.
[52]. Cho SH, Lee JS, Thabane L, Lee J. Acupuncture for obesity: a systematic review and meta-analysis. Int J Obes (Lond). 2009 Feb;33(2):183-96.
[53]. Du XZ, Bao CL, Dong GR, Yang XM. Immediate effects of scalp acupuncture with twirling reinforcing manipulation on hemiplegia following acute ischemic stroke: a hidden association study. Neural Regen Res. 2016 May;11(5):758-64.
[54]. Fragoso AP, Ferreira Ade S. Evaluation of the immediate effects of manual acupuncture on brachial bicep muscle function in healthy individuals and post stroke patients: a study protocol of a parallel-group randomized clinical trial. Zhong Xi Yi Jie He Xue Bao. 2012 Mar;10(3):303-9.
[55]. Du X, Wang J, Bao C, Jiao Z, Dong G. [Immediate effects of twirling reinforcing-reducing manipulation at head acupoints on muscle force in patients with acute ischemic stroke: a randomized controlled trial]. Zhongguo Zhen Jiu. 2016 Jan;36(1):43-7.
[56]. Plaster R, Vieira WB, Alencar FA, Nakano EY, Liebano RE. Immediate effects of electroacupuncture and manual acupuncture on pain, mobility and muscle strength in patients with knee osteoarthritis: a randomised controlled trial. Acupunct Med. 2014 Jun;32(3):236-41.
[57]. Lu TW, Wei IP, Liu YH, Hsu WC, Wang TM, Chang CF, Lin JG. Immediate effects of acupuncture on gait patterns in patients with knee osteoarthritis. Chin Med J (Engl). 2010 Jan 20;123(2):165-72.
[58]. Morais N, Greten HJ, Santos MJ, Machado JP. Immediate effects of acupuncture on the mechanosensitivity of the median nerve: an exploratory randomised trial. Acupunct Med. 2018 Jun;36(3):132-138.
[59]. Simma I, Gleditsch JM, Simma L, Piehslinger E. Immediate effects of microsystem acupuncture in patients with oromyofacial pain and craniomandibular disorders (CMD): a double-blind, placebo-controlled trial. Br Dent J. 2009 Dec 19;207(12):E26.
[60]. Irnich D, Behrens N, Gleditsch JM, Stör W, Schreiber MA, Schöps P, Vickers AJ, Beyer A. Immediate effects of dry needling and acupuncture at distant points in chronic neck pain: results of a randomized, double-blind, sham-controlled crossover trial. Pain. 2002 Sep;99(1-2):83-9.
[61]. Satoh H. Acute effects of acupuncture treatment with Baihui (GV20) on human arterial stiffness and wave reflection. J Acupunct Meridian Stud. 2009 Jun;2(2):130-4.
[62]. Matsumoto-Miyazaki J, Miyazaki N, Murata I, Yoshida G, Ushikoshi H, Ogura S, Minatoguchi S. Traditional Thermal Therapy with Indirect Moxibustion Decreases Renal Arterial Resistive Index in Patients with Chronic Kidney Disease. J Altern Complement Med. 2016 Apr;22(4):306-14.
[63]. Costa LA, de Araujo JE. The immediate effects of local and adjacent acupuncture on the tibialis anterior muscle: a human study. Chin Med. 2008 Dec 18;3:17.
[64]. Wang LL, Shan L, Du L, Zhang Y, Jia FY. Comparison of Electroacupuncture and Body Acupuncture on Gastrocnemius Muscle Tone in Children with Spastic Cerebral Palsy: A Single Blinded, Randomized Controlled Pilot Trial. Chin J Integr Med. 2020 Jan;26(1):14-19.
[65]. Ma YX, Ma LX, Liu XL, Ma YX, Lv K, Wang D, Liu JP, Xing JM, Cao HJ, Gao SZ, Zhu J. A comparative study on the immediate effects of electroacupuncture at Sanyinjiao (SP6), Xuanzhong (GB39) and a non-meridian point, on menstrual pain and uterine arterial blood flow, in primary dysmenorrhea patients. Pain Med. 2010 Oct;11(10):1564-75.
[66]. Akimoto T, Nakahori C, Aizawa K, Kimura F, Fukubayashi T, Kono I. Acupuncture and responses of immunologic and endocrine markers during competition. Med Sci Sports Exerc. 2003 Aug;35(8):1296-302.
[67]. Jones AY, Ngai SP, Hui-Chan CW, Yu HP. Acute Effects of Acu-TENS on FEV1 and Blood Β-endorphin Level in Chronic Obstructive Pulmonary Disease. Altern Ther Health Med. 2011 Sep-Oct;17(5):8-13.
[68]. Scheewe S, Vogt L, Minakawa S, Eichmann D, Welle S, Stachow R, Banzer W. Acupuncture in children and adolescents with bronchial asthma: a randomised controlled study. Complement Ther Med. 2011 Oct;19(5):239-46.
[69]. Ng DK, Chow PY, Ming SP, Hong SH, Lau S, Tse D, Kwong WK, Wong MF, Wong WH, Fu YM, Kwok KL, Li H, Ho JC. A double-blind, randomized, placebo-controlled trial of acupuncture for the treatment of childhood persistent allergic rhinitis. Pediatrics. 2004 Nov;114(5):1242-7.
[70]. Lang PM, Stoer J, Schober GM, Audette JF, Irnich D. Bilateral acupuncture analgesia observed by quantitative sensory testing in healthy volunteers. Anesth Analg. 2010 May 1;110(5):1448-56.
[71]. Liu S, Peng S, Hou X, Ke M, Chen JD. Transcutaneous electroacupuncture improves dyspeptic symptoms and increases high frequency heart rate variability in patients with functional dyspepsia. Neurogastroenterol Motil. 2008 Nov;20(11):1204-11.
[72]. Monnier L, Colette C, Owens D. La variabilité glycémique : l’évaluation de son impact passe par des index de mesure et des recommandations simples. Médecine des maladies Métaboliques. 2018;12(3):268-278.
[73]. Mori H, Okada Y, Kurozumi A, Narisawa M, Tanaka Y. Factors influencing inter-day glycemic variability in diabetic outpatients receiving insulin therapy. J Diabetes Investig. 2017;8(1):69-74.
[74]. Joshi A, Mitra A, Anjum N, Shrivastava N, Khadanga S, Pakhare A, Joshi R. Patterns of Glycemic Variability During a Diabetes Self-Management Educational Program. Med Sci (Bâle). 2019 25 mars 7(3):52.
[75]. Giacobino A, Petitpierre M, Dong HG. Acupuncture et stress : une contribution épigénétique. Acupuncture & Moxibustion. 2015;14(4):272-275